Note on Accessibility: Persons using mobile devices may find some tables are not fully accessible. Note that you can view tables on a larger screen or in the PDF version of the report/monograph. If you need additional assistance, email us or use our contact form and identify the tables for which access is required. We will assist you in accessing the content. NIH has helpful information on accessibility.
Toxicity Report 107
NTP Technical Report on the Toxicity Study of Stachybotrys chartarum (CASRN 67892-26-6) Administered by Inhalation to B6C3F1/N Mice

Abstract
Stachybotrys chartarum, also known as “black mold,” is a cellulolytic saprophyte with a worldwide distribution. Public concern for potential illnesses associated with water-damaged indoor environments has been heightened since the report of pediatric acute idiopathic pulmonary hemorrhage/hemosiderosis cases in the United States and following recent natural disasters. Although mycotoxicosis and pulmonary immunological endpoints have been previously examined, the systemic toxicity following subchronic inhalation of viable S. chartarum remains undercharacterized. To evaluate the toxicological responses to S. chartarum, B6C3F1/N mice were exposed to viable S. chartarum conidia (spores) twice a week for 3 months. All in-life procedures, including inhalation exposure, test article preparation, and hematology analysis, were completed by the National Institute for Occupational Safety and Health (NIOSH, Morgantown, WV). Battelle (Columbus, OH) conducted terminal necropsies, measured terminal body and organ weights, and evaluated gross lesions onsite at the NIOSH facility. Tissue processing and histopathology were completed at Battelle. Genetic toxicology studies on mouse peripheral blood erythrocytes were conducted by Integrated Laboratory Systems, LLC (Research Triangle Park, NC).
During the 3-month exposure study, groups of 10 male and 10 female B6C3F1/N mice repeatedly inhaled an estimated pulmonary dose of 1 × 104 viable S. chartarum conidia, 1 × 104 nonviable conidia (heat-inactivated particle control), or high-efficiency particulate air [filter] (HEPA)-filtered air. With the exception of one male mouse in the air control group, all mice survived to study termination. There was no effect of exposure on body weights. No gross lesions were observed at study termination. Mean absolute and relative lung weights were higher in male mice (11% and 5%, respectively) and significantly increased in female mice (33% and 35%, respectively) exposed to viable S. chartarum compared to the air control group.
Nonneoplastic lesions were observed in the larynx, lung, and bronchial lymph nodes. In males and females exposed to viable S. chartarum, nonneoplastic lesions were restricted mainly to the larynx and lung; however, there was a small incidence of lymphocyte hyperplasia in the bronchial lymph nodes of S. chartarum-exposed female mice (25%). In the larynx, epithelial squamous metaplasia was present at the base of the epiglottis in both males and females exposed to viable S. chartarum, whereas exposure to heat-inactivated particle control conidia did not significantly affect the larynx. The increased lung weights in the viable S. chartarum-exposed groups corresponded histopathologically with perivascular chronic-active inflammation, histiocytic cellular infiltration, medial hypertrophy of small- to medium-sized pulmonary arteries, goblet cell metaplasia, and bronchiolar epithelium hyperplasia in all exposed males and females. Lymphocyte hyperplasia of the bronchus-associated lymphoid tissue (BALT) was also present in the majority of viable S. chartarum-exposed males and females. Thrombi within pulmonary arteries were present in a limited number of S. chartarum-exposed males and females. The pulmonary histopathology of mice in the heat-inactivated particle control groups exhibited no lesions. Grocott’s methenamine silver (GMS)-stained lung sections of viable S. chartarum-exposed mice revealed conidia within the alveoli and alveolar macrophages. In the bronchial lymph nodes, lymphocyte hyperplasia and conidia were additionally observed in two viable S. chartarum-exposed female mice.
Under the conditions of this 3-month study, target organs identified in B6C3F1/N mice following inhalation exposure to viable S. chartarum conidia were the larynx, lung, and (to a lesser extent) the bronchial lymph nodes. The histopathological changes observed following viable S. chartarum exposure were not seen in the heat-inactivated particle control or air control mice. These results build on initial pulmonary immunology studies conducted at NIOSH using the same exposure parameters and demonstrate that the immunological responses and histopathology could be mediated by the viable S. chartarum bioaerosol.
Synonyms: Stachybotrys chartarum (S. chartarum); Stachybotrys atra (S. atra); Stachybotrys alternans (S. alternans); Stachybotrys chartarum (Ehrenb.) S. Hughes 1958; anamorphic, satratoxin-producing chemotype (CBS 414.95; IBT 9460)
Summary of Findings Considered Toxicologically Relevant in Male and Female Mice Exposed to Stachybotrys chartarum by Inhalation for Three Months
| Male Heat-inactivated Particle Control | Male Viable S. chartarum | Female Heat-inactivated Particle Control | Female Viable S. chartarum | |
|---|---|---|---|---|
| Exposure concentration (estimated lung deposition) | 1 × 104 heat-inactivated S. chartarum conidia | 1 × 104 viable S. chartarum conidia | 1 × 104 heat-inactivated S. chartarum conidia | 1 × 104 viable S. chartarum conidia |
| Survival rates | 10/10 | 10/10 | 10/10 | 10/10 |
| Body weights | No effect | No effect | No effect | No effect |
| Clinical findings | Nonea | None | None | None |
| Organ weights | None | None | None | ↑ Absolute and relative lung weights compared to either air control or heat-inactivated particle control |
| Hematology | None | ↑ Eosinophils compared to either air control or heat-inactivated particle control | None | None |
| Nonneoplastic effects | None | Larynx: epiglottis, metaplasia, squamous (4/10) Lung: perivascular, inflammation, chronic-active (10/10); artery, hypertrophy, medial (10/10); artery, thrombus (3/10); bronchiole, epithelium, hyperplasia (10/10); BALT, hyperplasia, lymphocyte (9/10); infiltration cellular, histiocyte (10/10); metaplasia, goblet cell (10/10) | Larynx: epiglottis, metaplasia, squamous (3/9) | Larynx: epiglottis, metaplasia, squamous (7/10) Lung: perivascular, inflammation, chronic-active (10/10); artery, hypertrophy, medial (10/10); artery, thrombus (1/10); bronchiole, epithelium, hyperplasia (10/10); BALT, hyperplasia, lymphocyte (9/10); infiltration cellular, histiocyte (10/10); metaplasia, goblet cell (10/10) Bronchial lymph node: hyperplasia, lymphocyte (2/8) |
| Genetic toxicology | ||||
| Micronucleated erythrocytes (in vivo): | Negative in males and females | |||
. Summary of Findings Considered Toxicologically Relevant in Male and Female Mice Exposed to Stachybotrys chartarum by Inhalation for Three Months
| Male Heat-inactivated Particle Control | Male Viable S. chartarum | Female Heat-inactivated Particle Control | Female Viable S. chartarum | |
|---|---|---|---|---|
| Exposure concentration (estimated lung deposition) | 1 × 104 heat-inactivated S. chartarum conidia | 1 × 104 viable S. chartarum conidia | 1 × 104 heat-inactivated S. chartarum conidia | 1 × 104 viable S. chartarum conidia |
| Survival rates | 10/10 | 10/10 | 10/10 | 10/10 |
| Body weights | No effect | No effect | No effect | No effect |
| Clinical findings | Nonea | None | None | None |
| Organ weights | None | None | None | ↑ Absolute and relative lung weights compared to either air control or heat-inactivated particle control |
| Hematology | None | ↑ Eosinophils compared to either air control or heat-inactivated particle control | None | None |
| Nonneoplastic effects | None | Larynx: epiglottis, metaplasia, squamous (4/10) Lung: perivascular, inflammation, chronic-active (10/10); artery, hypertrophy, medial (10/10); artery, thrombus (3/10); bronchiole, epithelium, hyperplasia (10/10); BALT, hyperplasia, lymphocyte (9/10); infiltration cellular, histiocyte (10/10); metaplasia, goblet cell (10/10) | Larynx: epiglottis, metaplasia, squamous (3/9) | Larynx: epiglottis, metaplasia, squamous (7/10) Lung: perivascular, inflammation, chronic-active (10/10); artery, hypertrophy, medial (10/10); artery, thrombus (1/10); bronchiole, epithelium, hyperplasia (10/10); BALT, hyperplasia, lymphocyte (9/10); infiltration cellular, histiocyte (10/10); metaplasia, goblet cell (10/10) Bronchial lymph node: hyperplasia, lymphocyte (2/8) |
| Genetic toxicology | ||||
| Micronucleated erythrocytes (in vivo): | Negative in males and females | |||
Introduction
Fungi, Indoor Environmental Quality, and Public Health
Fungi are a diverse kingdom with an estimated 5.1 million species.3 These eukaryotic organisms contain membrane-bound organelles with a rigid cell wall composed of mannose proteins, complex polysaccharides such as α/β glucans, chitin, and ergosterol.4,5 The lifecycles of fungi can be unicellular yeast, multicellular hyphae, or a combination of the two (dimorphism). Fungi reproduce asexually or sexually and produce spores, which can also be produced by a variety of other organisms including plants and bacteria. Conidia are asexual fungal spores that are morphologically distinct, ranging from 2 to >120 µm in size.4 Fungi are mostly saprophytic heterotrophs that obtain nutrients from decaying organic material, allowing growth in environments ranging from soil, agricultural commodities, and indoor environments containing cellulose-based building material. Although most fungal species are not pathogenic, some cause disease in plants, animals, and humans, and the potential for adverse health effects following fungal exposure has become an area of public concern in the United States.
Damp indoor environments may provide optimal growth conditions required for fungal proliferation. Abiotic or biotic disturbances can result in the aerosolization of fungal bioaerosols composed of spores, filamentous hyphae, or microscopic fragments into the surrounding environment. Exposure can also occur in occupational settings, including forestry, agriculture, service, and manufacturing, and can consist of spore concentrations exceeding 108 colony-forming units per cubic meter (CFU/m3).6 Along with spores, hyphal fragments can also be aerosolized into the breathing zone of exposed individuals and can be found at higher concentrations than spores.7-12 Fungal bioaerosols can contain secondary microbial metabolites, such as mycotoxins and microbial volatile organic compounds (MVOCs), that have the potential to further exacerbate the negative health responses following exposure.
A study of water-damaged building materials collected from damp indoor environments (n = 72) identified fungal genera including Penicillium (68%), Aspergillus (56%), Chaetomium (22%), Ulocladium (21%), Stachybotrys (19%), and Cladosporium (15%), all of which are known to cause respiratory allergy.13 Of these six commonly identified fungi, Stachybotrys chartarum was one of the most frequently detected fungal species. In 1994, evidence of acute idiopathic pulmonary hemosiderosis (IPH) was identified in infants residing in Cleveland, OH.14,15 Further investigation suggested that more of the infants diagnosed with acute IPH resided in water-damaged homes than in homes that did not contain water damage, and S. chartarum was identified as a potential causative agent.15 However, a later report published by the Centers for Disease Control and Prevention refuted those results, stating that the evidence presented in the initial report did not support the epidemiological associations between water damage, S. chartarum exposure, and acute IPH.15 Although the initial association between human illness and mold contamination was nullified, awareness of the association between fungal exposure and human health was already heightened.
Following the Cleveland investigation, consensus documents from the Institute of Medicine, Committee on Damp Indoor Spaces and Health,16 and the World Health Organization17 identified epidemiological evidence of associations between exposure to mold contamination in damp indoor environments and adverse respiratory symptoms such as asthma, dyspnea, wheeze, cough, respiratory infections, bronchitis, allergic rhinitis, eczema, and upper respiratory tract symptoms.18 In addition, fungal exposure has been associated with allergic bronchopulmonary mycoses and hypersensitivity pneumonitis.6,19 In 2017, cost estimate analyses indicated that the direct health-care costs of all fungal-related diseases exceeded $7.2 billion, which included $4.5 billion from hospitalizations and $2.6 billion from outpatient visits.20 Although associations between mold contamination in damp indoor environments and adverse health effects have been identified, the pulmonary and systemic toxicity resulting from repeated fungal exposure remains largely unknown. In addition, there are no current exposure limits or regulations in place to protect individuals who encounter contaminated fungal environments.
To address these knowledge gaps, fungi were nominated to the National Toxicology Program (NTP) in May 2004. The NTP Board of Scientific Counselors and Executive Committee suggested studying fungal species commonly identified in indoor environments, including Aspergillus, Alternaria, and Stachybotrys. In 2007, an expert review panel discussed the NTP subchronic study design, including the fungal test article production, characterization, and reproducibility of the test article generation. As a participant of the review panel, the National Institute for Occupational Safety and Health (NIOSH) was identified as a collaborative partner to conduct subchronic (13-week) inhalation studies to examine the toxicological effects following repeated fungal exposure. To complete these subchronic inhalation exposures, NIOSH developed a computer-controlled acoustical generator system (AGS) to deliver dry aerosolized fungal test articles to mice housed in nose-only chambers to model a natural human inhalation exposure that would occur in residential or occupational environments. The use of the AGS circumvents limitations of other administration methods, including intranasal exposure,21 intratracheal instillation,22-25 and liquid aerosol inhalation.26 Environmentally and occupationally relevant fungal species nominated to NTP have been independently tested in both pilot immunology studies conducted by NIOSH27-29 and in toxicology studies reported by NTP in collaboration with NIOSH.30
A 4-week and 13-week pilot exposure study using the AGS evaluated the pulmonary immune responses to repeated S. chartarum exposure.27 Following the initial optimization and characterization of the S. chartarum test articles, including a heat-inactivated biological particle control,28 groups of B6C3F1/N female mice repeatedly inhaled one of two viable trichothecene-producing strains of S. chartarum twice a week for a total of 4 or 13 weeks.27 The two S. chartarum strains used in the pilot study were IBT 9460 and IBT 7711. IBT 9460 is a macrocyclic trichothecene-producing strain that produced a higher level of mycotoxin and fragmented to a greater extent than IBT 7711.28 Separate groups of mice inhaled either nonviable S. chartarum conidia (a heat-inactivated particle control for each respective strain) or high-efficiency particulate air [filter] (HEPA)-filtered air only. Pulmonary arterial remodeling, characterized by airway inflammation with a granulocyte infiltration, was observed after 4 weeks of S. chartarum IBT 9460 exposure. Granulocyte infiltration switched to more of a lymphocytic and macrophage infiltration after 13 weeks of exposure to both IBT 9460 and IBT 7711. Perivascular inflammation and bronchiolar epithelial cell hyperplasia were also observed. Flow cytometry analysis of bronchoalveolar lavage fluid (BALF) from exposed mice showed an increase in eosinophils, neutrophils, and macrophages following a 13-week exposure to both IBT 9460 and IBT 7711. However, the progression of cellular infiltrates and immunopathological responses progressed more rapidly in IBT 9460-exposed mice compared to IBT 7711-exposed mice, suggesting that the presence of fungal fragments plays a critical role in the pulmonary immune response to S. chartarum exposure.
In an NTP study conducted in collaboration with NIOSH describing the toxicological responses following a subchronic A. fumigatus exposure, the incidence of nonneoplastic lesions was significantly increased in the larynx, lung, and bronchial lymph nodes of viable A. fumigatus-exposed male and female B6C3F1/N mice compared to the control groups.30 In the lung, arterial medial hypertrophy was identified and described as circumferential thickening of the tunica media of small and medium arterioles and arteries by concentric layers of hypertrophied and hyperplastic smooth muscle cells. In addition, chronic-active inflammation, defined as the increased presence of neutrophils, was observed in the lung tissue following subchronic exposure to A. fumigatus. Bronchus-associated lymphoid tissue (BALT) hyperplasia and goblet cell metaplasia were also observed following A. fumigatus exposure. Squamous metaplasia of the epiglottis was observed in the larynx of both male and female mice exposed to viable A. fumigatus compared to the heat-inactivated particle control and air control groups. Evaluation of the bronchial lymph nodes showed significantly increased incidences of lymphocyte hyperplasia and plasma cell hyperplasia relative to either the heat-inactivated particle control or air control groups.30
The purpose of these studies on A. fumigatus was to characterize the toxicological and pulmonary immunological responses following repeated exposure to fungal contaminants commonly identified in occupational and damp indoor environments. Using a method that replicates a more natural human exposure has provided unique opportunities to define the mechanisms by which these fungal species cause adverse respiratory health effects. To further understand the hazards following S. chartarum exposure, NIOSH employed this same methodology to characterize the toxicological responses following subchronic exposure to S. chartarum strain IBT 9460, a high trichothecene-producing, high fragmenting strain of S. chartarum.
Chemical and Physical Properties
The genus Stachybotrys is an asexually reproducing, filamentous fungal genus that includes 50 species found worldwide and belongs to the Ascomycota order Hypocreales, family Stachybotryaceae.31 S. chartarum is a macroscopic saprophyte that is greenish black in color, leading to the common name “black mold.” This fungus was first identified in 1818 as Stillbospora chartarum Ehrenb.32 In 1837, August Carl Joseph Corda identified the fungus as Stachybotrys atra following collection from wallpaper in a house in Prague,33 which was then reclassified as S. chartarum (Ehrenb.) S. Hughes 1958.34 S. chartarum requires a cellulose-rich substrate, such as drywall, wallpaper, gypsum board, or cardboard, and high moisture content for optimal growth.35-37 S. chartarum can grow at a range of temperatures, but the most optimal condition for growth is 30°C at 93%–94% humidity.38 Frazer and colleagues39 showed different temperature and water activities are required for the germination and growth of S. chartarum compared to sporulation. S. chartarum is often found in damp indoor environments.1,11,40-42 Consequently, one study comparing water-damaged building materials collected from damp indoor environments (n = 72) identified S. chartarum more often than other fungal species such as Penicillium chrysogenum and A. versicolor.13
The mycelium of S. chartarum is characterized by the formation of dark, slimy masses of conidia (spores) that sporulate abundantly from phialides. The phialides are 9–14 mm in length and often occur in whorls that extend from the tip of conidiophores connected to septate hyphae. S. chartarum conidia are smooth or ridged, ellipsoid, and unicellular and range from 7 to 12 µm in length and 4 to 6 µm in width.43 The aerodynamic diameter of aerosolized S. chartarum (strain IBT 9460) conidia has been reported to be approximately 3–5 µm28; however, one study showed that North American strains have an average aerodynamic diameter of 5.6 µm, but the conidia size was variable, and approximately one-third of the conidia tested were within the respiratory range.43 This high variability in conidia size could potentially be due to the presence of fungal fragments, a notion supported by aerosolization studies describing the high occurrence of fungal fragments. One study conducted by Lemons et al. used S. chartarum strain IBT 9460 and showed that acoustical generation resulted in the aerosolization of not only intact S. chartarum conidia (3–5 µm) but also of fungal fragments (<2 µm aerodynamic diameter) derived from conidia, phialides, and hyphae that initially constituted 50% of the total fungal particle count.28 Cho et al. conducted a separate study, which found that S. chartarum fungal fragments were aerosolized at concentrations 514 times higher than S. chartarum spores.8
It has been suggested that the mycotoxins produced by S. chartarum contribute to the adverse health effects observed following exposure.22,37,44-46 Two chemotypes of S. chartarum exist, chemotype S and chemotype A, and are defined by the mycotoxins produced. Used for this current study, chemotype S produces toxic macrocyclic trichothecenes including satratoxins, roridins, and verrucarins,47,48 whereas chemotype A produces less toxic atranones.1,49 In addition, S. chartarum also produces phenylspirodrimanes,41 trichoverrols,50 trichoverrins,50 and spirocyclic drimanes,1 as well as MVOCs,51 which individuals could encounter within S. chartarum-contaminated environments.
Human Exposure
S. chartarum is distributed worldwide and is found in soil and on cellulose-rich materials including hay, straw, plant debris, and building materials.35,37,52 Human exposure to contaminated materials, such as straw contaminated with S. chartarum, has resulted in dermatological and adverse respiratory health effects.53,54 Exposure to S. chartarum can occur via dermal contact, ingestion, and inhalation; however, studies have shown that exposure occurs primarily through direct skin contact and inhalation.44,54 S. chartarum spores do not readily disperse into the air because of the slime droplet encasing the phialides but can become aerosolized when the fungus dries and is disturbed via biotic or abiotic air movements.44 Exposure to fungal bioaerosols, which include S. chartarum, generated from disturbances within fungal-contaminated, water-damaged environments have been associated with poor indoor air quality11,48 and adverse respiratory health effects.16,17
Fungal bioaerosols can be composed of spores, filamentous hyphae, and microscopic fragments. Fungal fragments have the potential to aerosolize at higher concentrations than spores.7-12 In addition, certain strains of S. chartarum fragment to a greater extent compared to other S. chartarum strains or fungal species11,28 and may contribute to the adverse respiratory outcomes following S. chartarum exposure.27 Case reports have identified an association between indoor S. chartarum exposures and respiratory symptoms,55-57 immunological dysfunction,58 and neurological disease; however, there is a lack of evidence for neurotoxicity.41
S. chartarum has been detected in water-damaged environments such as schools, offices, and home dwellings13,58-61 and has been associated with “sick building syndrome” and damp-related building illness.44,46,59,62-66 Workers remediating S. chartarum-contaminated material from buildings reported symptoms, including throat irritation, fatigue, muscle aches, and skin rashes.57 Sampling of contaminated work areas resulted in an estimated air concentration of 2 × 104 CFU/m3,58,67 which coincided with other measured S. chartarum airborne levels reported to be 7 × 105 viable conidia/m3.43 A case control study of 10 infants with pulmonary hemorrhage from Cleveland, OH reported the mean air concentration of S. atra (synonym S. chartarum) was 43 CFU/m3 compared to 4 CFU/m3 in the homes of infants without pulmonary hemorrhage.68 Residents living in water-damaged homes have shown severe asthma with an allergic reaction to Stachybotrys antigens isolated from moldy carpets.69 Neurological and flu-like symptoms, fatigue, dermatitis, and malaise have also been reported from a home with identified Stachybotrys contamination, symptoms of which resolved following the removal of Stachybotrys-contaminated material.55 Several studies that have characterized the responses of individuals exposed to S. chartarum suggested that the symptoms reported were likely caused by inhalation of the fungal toxins.56,58,61
S. chartarum is considered nonpathogenic.70 The ability of the species to contribute to adverse health outcomes is suggested to be caused by the allergenicity, inflammatory response, or toxicity of S. chartarum. It was once thought that S. chartarum was incapable of infection because of the lack of physical evidence of the organism within a symptomatic individual. Most recently, however, Stachybotrys (S. chlorohalonata) was identified in the sinus tissues of a severely immunocompromised 23-year-old man.71 Although mycotoxins were also reported to contribute to the hemorrhaging observed in the immunocompromised male patient, this was the first report of an invasive case of Stachybotrys.
Regulatory Status
Currently, there are no established guidelines for acceptable Stachybotrys concentrations in residential or occupational environments. In addition, no exposure limits have been proposed by any regulatory or governmental body.
Toxicity
S. chartarum has been characterized as a toxigenic mold that can cause animal and human mycotoxicosis.41 The effect of S. chartarum trichothecenes was identified in the 1930s Russian equine outbreak when horses died after consuming S. chartarum-contaminated straw and grain.52 This newly found disease caused by ingesting or handling contaminated feed was termed stachybotryotoxicosis and has been reported worldwide in animals and humans.43,53,72 Farm workers exposed to S. chartarum-contaminated hay complained of dyspnea, airway obstruction, conjunctivitis, sore throat, mucosal irritation, and skin inflammation.73 Previous studies suggest that the occupational exposure concentrations resulting in trichothecene toxicosis were on the order of 103–107 conidia/m3.43 Although these mycotoxicosis cases were due to direct skin contact or ingestion, inhaling mycotoxins is considered more harmful because of the greater fungal bioavailability.43
Distinct mycotoxin profiles are produced by S. chartarum when grown on different culture media.74 However, research has indicated that the water activity (aw) must reach a level of 0.95 aw before mycotoxins are produced.48 S. chartarum produces macrocyclic trichothecenes, such as satratoxins and isosatratoxins F, G, and H, roridin E and L-2, and verrucarins J and B. These macrocyclic trichothecenes are suggested to contribute in part to the adverse health effects following S. chartarum exposure.22,37,44-48 Research has shown that S. chartarum macrocyclic trichothecenes within indoor contaminated environments can become airborne and are associated with both conidia75 and fungal fragments.76,77 Mycotoxins have been associated with particle sizes ≥3 µm, which correspond to conidia; however, trichothecene mycotoxins, such as satratoxin H and verrucarin J, have been shown to be associated with other S. chartarum components, including submicron fragments that can be inhaled and deposited deep within the lung.78
In general, satratoxins have been reported to be produced in larger amounts compared to the other trichothecenes.72 In mice, the median lethal dose (LD50) for satratoxin has been reported to be approximately 1 mg/kg.79 Following the aerosolization of S. atra (syn. S. chartarum) conidia, Sorenson et al. confirmed the presence of satratoxin H within the bioaerosol, as well as satratoxin G and trichoverrols A and B to a lesser extent.80 Satratoxin G has been reported to be the most cytotoxic of the satratoxins and five other trichothecenes studied.81 A single dose of ≥25 µg/kg satratoxin G administered via intranasal instillation resulted in neurotoxicity and inflammation within the nose and brain of female C57Bl/6 mice.82 Isosatratoxin F exposure altered the phospholipid synthesis, composition, and secretion of pulmonary surfactant in BALF in mice83 and resulted in cytological alterations within fetal rabbit alveolar type II cells, indicative of cellular damage and apoptosis. Additionally, satratoxin-positive S. chartarum, as well as the trichothecene mycotoxins (roridin A, verrucarin A, and T-2 toxin), have been shown to induce apoptosis and inflammation in human primary macrophages.84
Stachylysin, a hemolysin released from S. chartarum,85-87 was thought to contribute to the infant pulmonary hemosiderosis cases reported in Cleveland, OH.14,15,88 To investigate this potential contribution, a study conducted by Vesper and Vesper88 measured the production of stachylysin from the S. chartarum strains isolated from infant pulmonary hemosiderosis case houses in Cleveland, OH46 and in an isolate from the lung of a pulmonary hemosiderosis patient in Texas.89 Results showed the isolated S. chartarum strains produced stachylysin, suggesting a potential role in the reported pulmonary hemosiderosis cases.88
Exposure to atranones A and C has also been shown to result in immunotoxic and inflammatory responses in the lungs of male Swiss Webster (CFW) mice following a single dose of ≥2 µg/animal via intratracheal instillation.90 Rand et al. suggested that different atranones exhibit varying levels of inflammation with different toxicokinetics.90 Another study showed that a single exposure to a nontoxin-producing strain of S. chartarum could induce airway irritation and elicit an immune response in male Swiss Webster (CFW) mice.26
Fungi also produce MVOCs, and similar to mycotoxins, the production of MVOCs depends on the growth media. For example, MVOCs unique to S. chartarum include 1-butanol, 3-methyl-2-butanol, 3-methyl-1-butanol, and thujopsene and have been detected on rice inoculated with S. chartarum, while 1-butanol was detected on inoculated gypsum board.51 MVOCs emitted from contaminated fungal environments have additionally been implicated in central nervous system symptoms such as headaches, inability to concentrate, or dizziness.91
Research surrounding the toxic effect of exposure to spores, secondary metabolites, or MVOCs of S. chartarum has been confounding. It has been predicted that the adverse health effects observed in animal models are caused by exposure to high spore or mycotoxin concentrations that exceed those encountered in even the most heavily contaminated environments.37,92 However, the pediatric IPH cases identified in Cleveland, OH15 occurred after exposure to fungal-contaminated environments, suggesting that the spore or mycotoxin concentrations encountered in these damp indoor environments were sufficient to cause adverse health effects in children. Additionally, the length of fungal growth and substrate availability influenced the secondary metabolite profiles and the associated effect on adverse respiratory health effects.93 Regardless of the variability in toxin production, the results of most toxicological studies demonstrate that S. chartarum produces highly toxic trichothecenes that have been shown to contribute to adverse health effects.
Reproductive and Developmental Toxicity
While not well studied, one report found that CFW male and female mice orally exposed to a single, low dose (3,000–4,000 tissue culture units) of S. alternans (syn. S. chartarum) prepared from grain or liquid medium infected with toxicogenic and nontoxicogenic strains resulted in a decreased number of pregnancies compared to control mice.94 In addition, mice from that same study that had ingested 100–4,000 tissue culture units of toxin had a significant increase in the frequency of dead, resorbed, or stunted fetuses compared to controls.
Carcinogenicity
Several studies have evaluated the carcinogenicity of individual mycotoxins; however, no studies evaluating carcinogenicity following S. chartarum exposure were identified.
Genetic Toxicity
Although studies evaluating the effect of purified mycotoxin exposure on genotoxicity exist, no studies evaluating the genetic toxicity following S. chartarum exposure were identified.
Study Rationale
Following the nomination of fungi to NTP for comprehensive toxicity evaluation, NTP partnered with NIOSH to begin a series of studies to characterize the toxicological responses following exposure to NTP-nominated fungal species. The first of the series examined the toxicological responses to A. fumigatus30 following delivery of dry aerosolized fungal test articles twice a day for 13 weeks to male and female B6C3F1/N mice housed in nose-only chambers. The design of this inhalation study was to simulate a more natural exposure similar to what would be encountered in a water-damaged built environment. In this second inhalation toxicity study, S. chartarum was selected as the test article because of the heightened awareness of adverse health effects of fungal exposures encountered within indoor water-damaged environments. Although pulmonary immunological endpoints have been evaluated in pilot studies, the toxicological responses to S. chartarum exposure have not been fully characterized.
Materials and Methods
Procurement and Characterization
Culture of Stachybotrys chartarum
Stachybotrys chartarum (CBS 414.95, IBT 9460) was originally isolated from a water-damaged building in Finland.1 This is a macrocyclic trichothecene-producing strain and can fragment to a greater extent than other S. chartarum strains.28 Bulk S. chartarum cultures were cultivated from 25% glycerol stocks prepared as described in Appendix A. Aliquots of the glycerol stock were stored at −80°C for the study duration.
Individual glycerol stock aliquots were used weekly to inoculate two malt extract agar (MEA) plates for the cultivation of S. chartarum according to the methods detailed in Appendix A. After 12–14 days of culture, S. chartarum conidia (spores) were liberated from each plate using 2 mL of sterile water. Twelve culture flasks containing wet, autoclaved white rice (30 g) were inoculated with 1.25 × 106 conidia. Following 21–28 days of growth, half of the flasks were heat-treated at 80°C for 2 hours to produce the heat-inactivated particle control. The remaining flasks served as the viable test article. The heat-inactivated particle control and viable S. chartarum flasks were transferred to the National Institute for Occupational Safety and Health (NIOSH) inhalation facility and placed in a desiccator for 7–10 days prior to aerosolization.
Quality Assurance
Quality assurance and quality control of heat-inactivated and viable S. chartarum consisted of testing cultures each week using internal transcribed spacer gene sequencing. This method allowed the amplification and detection of potentially contaminating mycobiota sources as described in Appendix A. Cultures were also tested weekly to ensure reproducibility and production of nonviable S. chartarum. All identified fungal DNA belonged to S. chartarum with no contaminating fungi detected in the viable or heat-inactivated S. chartarum cultures. Heat inactivation resulted in an average of 98.63% reduction in S. chartarum (Table A-1).
Test Article Generation and Exposure System
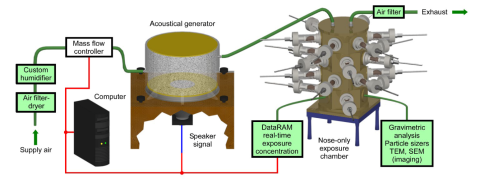
NIOSH has developed a computer-controlled acoustical generator system (AGS), in which acoustical energy is used to release the fungal spores into a nose-only exposure chamber. Figure 2 shows the AGS based on a modified acoustically powered particle (Pitt-3) generator, which was originally described by Weyel et al.95 The Pitt-3 generator was purchased from the University of Pittsburgh and evaluated under conditions recommended by its engineering school.96 The system was then fitted with a computer controller, further modified to its current design,97,98 and used by NIOSH to study the respiratory effects caused by the inhalation of a wide variety of occupationally relevant dusts. The AGS was initially used as a new and improved system for dispersing nanomaterials99 and was subsequently modified for fungal spore aerosolization as described by Buskirk et al.100 Fungi were grown on rice as described above, placed on the rubber membrane of the AGS, and aerosolized using acoustical energy. Inhalation of fungal spores simulates natural fungal exposures in the environment better than other routes of exposure. Several studies conducted by NIOSH to date using the AGS demonstrated that fungal aerosols can be reproducibly administered to mice housed in the nose-only exposure chamber,27-29,100-102 including a National Toxicology Program (NTP) study assessing the toxicological response to Aspergillus fumigatus.30
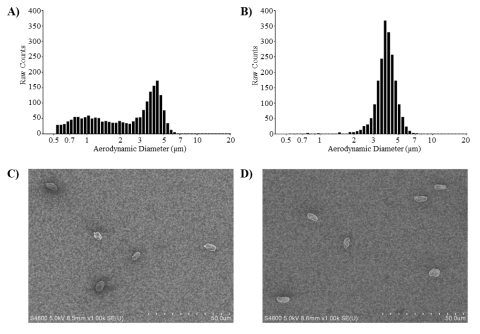
Initial aerosolization studies demonstrated that the AGS produced an S. chartarum aerosol composed of single conidia of similar size (3–5 µm) and morphology in both the viable test article (S. chartarum) and heat-inactivated particle control groups (Figure 3). However, unlike the heat-inactivated particle control, a significant proportion of the viable S. chartarum aerosol was also composed of smaller fragments (<2 µm, Figure 3A).28
Aerosolized viable S. chartarum and the heat-inactivated particle control were delivered to male and female B6C3F1/N mice housed in a multianimal nose-only exposure chamber via the AGS as previously described.27,28 Animals were exposed two times per week for 3 months via inhalation. In addition, an air control group received the same high-efficiency particulate air [filter] (HEPA)-filtered air that was being used to deliver viable S. chartarum.
Inhalation exposures were achieved by weighing and then placing the mice in a custom-built nose-only multianimal exposure carousel connected to the AGS as described in McKinney et al.99 Mice were placed in random pods for each exposure to minimize the effects of slight concentration differences observed within individual ports. Although the nose-only carousel houses up to 24 pods (Figure 2), only 20 pods (10 male and 10 female mice) were used during each exposure. Additional ports served as sample ports for the collection and analysis of test articles. The air control, heat-inactivated particle control, and viable S. chartarum-exposed animals were exposed using different chambers located in the same inhalation exposure room.
B6C3F1/N mice were exposed to viable S. chartarum or heat-inactivated particle control until the DataRAM software estimated that a total of approximately 1 × 104 conidia had been deposited within the lungs during the 1-hour exposure session. Figure 4 shows the estimated pulmonary deposition following each exposure for the duration of the study. The estimation software used a mouse-lung-deposition model developed by the NIOSH Inhalation Facility, which was based on deposition measurements from Raabe et al.103 During viable S. chartarum or heat-inactivated particle control exposures, lung-deposition calculations were achieved using real-time DataRAM mass concentration measurements and a previously described scaling factor that estimated the number of conidia deposited in the lungs of mice in real time.100,102 The integrated value of mass deposition per unit time was calculated throughout the exposures. The AGS automatically turned off when the desired total conidia deposition had been delivered to the mice; however, the mice continued to receive HEPA-filtered humidified (approximately 40% relative humidity) air supplied at a flow rate of 6 L/min until 60 minutes had been reached. Each exposure corresponded to a 60-minute average mass concentration of approximately 8 mg/m3 (viable S. chartarum) or 7 mg/m3 (heat-inactivated particle control). Gravimetric filters were also collected during each exposure to continually monitor and adjust the DataRAM calibration. Air control mice received only HEPA-filtered air supplied to the chamber at the same flow rate (6 L/min) and time (60 minutes) as S. chartarum-exposed mice. All environmental conditions were kept constant in each carousel between control and test article animals.
Animal Source
Male and female B6C3F1/N mice (n = 20/exposure group and 20 sentinel mice) were obtained from the NTP colony maintained by Taconic Biosciences, Inc. (Germantown, NY).
Animal Welfare
Animal care and use were in accordance with the Public Health Service Policy on Humane Care and Use of Animals. The NIOSH Inhalation Facility is an environmentally controlled barrier facility that is fully accredited by AAALAC International. Studies were approved by the CDC-Morgantown Animal Care and Use Committee and conducted in accordance with all NIOSH animal procedures and policies.
Exposure Concentration Selection Rationale
An estimated pulmonary dose of 1 × 104S. chartarum conidia (viable or heat-inactivated conidia) was chosen for this study. This concentration was the highest aerosolized dose that reproducibly induced a lymphoproliferative response and was the same concentration and schedule used for a previously published pulmonary immunology study.27,28 This estimated lung burden would be equivalent to that of a worker breathing a constant workplace air concentration of 3.45 × 106 conidia/m3 (146 μg/m3) for 1 week, working 8 hours per day for 5 days, and is comparable to an occupational exposure concentration of 103–107 conidia/m3, which has been reported to result in toxicosis.43
Three-month Study
Mice were approximately 5 to 6 weeks old on receipt. The animals were weighed, distributed randomly into groups of approximately equal initial mean body weights, and randomly assigned to viable S. chartarum, heat-inactivated particle control, or air control groups. Mice were initially identified via tail marking, using a permanent marker and a consecutive numbering system.
Animals that weighed >10% or <10% of the mean of the group were placed in the sentinel group (n = 20). At 4 weeks and at study termination, serological analyses for bacterial and viral pathogens were performed on 10 male and 10 female sentinel mice using the protocol of the NIOSH Sentinel Animal Program (Appendix C).
Before the study, the mice were acclimated to the NIOSH animal facility for a minimum of 7 days. Afterward, to reduce potential stress from exposures, the mice were acclimated to the housing units used for the exposures (nose-only carousel) for 1 week (two 1-hour acclimations) before the initial fungal exposure. Study mice were placed individually in the carousel housing units (pods), and humidified HEPA-filtered air was supplied to the chamber for 1 hour on 2 separate days during the week before the initial exposures. After acclimation was complete and before the exposures began, the mice were tattooed using a consecutive numbering system.
Male and female B6C3F1/N mice were randomly assigned to viable S. chartarum, heat-inactivated particle control, and air control groups. Using a modified NTP protocol, groups of 10 male and 10 female mice were placed in individual pods on the carousel on the same 2 days of the week for 26 exposures (3 months). The mouse positions in the carousel were rotated throughout the study to account for any dose variability based on pod placement. Male and female mice were exposed for 1 hour per day to 8 mg/m3 (resulting lung dose = 1 × 104) viable S. chartarum conidia, 7 mg/m3 (resulting lung dose = 1 × 104) heat-inactivated S. chartarum conidia (particle control), or HEPA-filtered air (air control) that was delivered through the AGS into the nose-only exposure chamber at a flow rate of 6 L/min.
Following acclimation and exposures, the animals were transferred from the individual carousel housing pods to a standard cage environment located in an adjoining room within the facility. Male and female mice were housed in HEPA-filtered ventilated polycarbonate cages on autoclaved hardwood chip bedding. Male B6C3F1/N mice were individually housed during acclimation and the in-life phases of the study because of social incompatibilities, whereas female mice were housed in groups of five. Enrichment was provided to all cages and consisted of crinkle cut kraft paper. The room temperature of the animal facility was maintained between 68°F and 76°F, and the relative humidity ranged between 17% and 70% (Table 1). The light/dark cycle was maintained at 12-hour intervals. Mice were provided feed and tap water ad libitum, except when housed in the pods during acclimation or exposures. Sentinel mice selected during the acclimation period were housed within the same room and were free of viral and bacterial pathogens (Appendix C).
Study animals were weighed upon initial receipt, and body weights were recorded weekly until study termination. Clinical observations were recorded daily for the duration of the study, once a day (morning) on nonexposure days and twice a day (morning/afternoon) on exposure days. Information about the study design, facilities, and animal maintenance is provided in Table 1. The report on feed composition and contaminants is provided in Appendix B.
Clinical Examinations and Pathology
At the completion of the 3-month study, the mice were injected intraperitoneally with a lethal dose of sodium pentobarbital euthanasia solution (100–300 mg/kg). Peripheral blood for hematology and micronuclei determination was obtained via cardiac puncture. Whole blood (a minimum of 600 µL) was collected in tubes containing potassium ethylenediaminetetraacetic acid, and 400 µL were immediately processed for hematology using a ProCyte Dx hematology analyzer (IDEXX Laboratories, Inc., Westbrook, ME). Table 1 lists the parameters measured. A minimum of 200 µL of blood was immediately refrigerated and shipped to Integrated Laboratory Systems, LLC (ILS; Research Triangle Park, NC) for micronuclei determination.
Necropsies were performed at the NIOSH site by Battelle staff (Columbus, OH) in random order on all animals surviving to terminal sacrifice. The organs weighed at terminal necropsy included the left and right epididymides, heart, left and right kidneys, liver, lungs, left and right ovaries, left and right testes, and thymus. Bilateral organs were weighed and recorded separately. At necropsy, the tissues in Table 1 were examined in situ for gross lesions, removed, and then fixed and preserved in 10% neutral buffered formalin. Exceptions included the eyes and testes (epididymis and vaginal tunics of testes), which were initially fixed in Davidson’s solution and modified Davidson’s solution, respectively, then transferred to 10% neutral buffered formalin. The collected and fixed tissues were then shipped to the Battelle facility and further processed and trimmed, embedded in paraffin, sectioned at a thickness of 5 µm, and stained with hematoxylin and eosin (H&E). Histological sections of aorta, gallbladder, heart, liver, lung (all five lobes), bronchial lymph node, and spleen were stained with Grocott’s methenamine silver (GMS) stain for fungal elements. Battelle laboratory pathologists conducted a complete histopathological examination on all air control, heat-inactivated particle control, and viable S. chartarum-exposed mice. Table 1 lists the tissues and organs that were examined.
Microscopic evaluations were completed by a board-certified veterinary pathologist, and the pathology data were entered into the NTP Provantis software (Instem Stone, UK). The report, slides, paraffin blocks, residual wet tissues, and pathology data were sent to the NTP Archives for inventory and storage. An audit of pathology specimens was conducted wherein the wet tissues, blocks, and slides were examined by technical staff for quality and adherence to NTP Specifications,104 and the wet tissues were examined by a team of pathologists to ensure all were sampled according to NTP Specifications. The slide and tissue counts were also verified. Slide-mounted, H&E-stained slides were evaluated by a team of quality assessment (QA) pathologists at a pathology laboratory independent of the study laboratory for accuracy and consistency of diagnoses. The histotechnique was also evaluated.
After a review of the laboratory reports and selected histopathology slides by a QA pathologist, the findings and reviewed slides were submitted to the Pathology Working Group (PWG) coordinator. Any inconsistencies in the diagnoses made by the study laboratory and QA pathologist were resolved by the DTT pathology peer-review process. Final diagnoses for reviewed lesions represent a consensus of the PWG or a consensus between the study laboratory pathologist, DTT pathologist, and QA pathologist/PWG coordinator. Details of these review procedures have been described, in part, by Boorman105 and Maronpot.106
Statistical Methods
Calculation and Analysis of Nonneoplastic Lesion Incidences
The incidences of nonneoplastic lesions are presented as numbers of animals bearing such lesions at a specific anatomic site and the numbers of animals with that site examined microscopically. Fisher’s one-sided exact test,107 a procedure that uses the overall proportion of affected animals, was used to determine significance between all pairs of the three exposure groups.
Analysis of Continuous Variables
Two approaches were employed to assess the significance of all pairwise comparisons between exposed and control groups in the analysis of continuous variables. Organ and body weight data, which historically have approximately normal distributions, were analyzed using a one-way analysis of variance with post hoc Tukey Honest Significant Difference test108 for all pairwise comparisons among the three exposure groups. Hematology data, which have typically skewed distributions, were analyzed using the nonparametric multiple comparison method of Dunn.109 Prior to statistical analysis, extreme values identified by the outlier test of Dixon and Massey110 were examined by DTT personnel, and implausible values were eliminated from the analyses.
Quality Management
The 3-month study was conducted in compliance with U.S. Food and Drug Administration Good Laboratory Practice Regulations whenever possible.111 To comply with the NIOSH Health Effects Laboratory Division (HELD) Laboratory Quality Management System, this study was conducted using validated standard operating procedures and performed in accordance with a CDC-Morgantown Animal Care and Use Committee-approved animal protocol, as well as a HELD Office of the Director-approved study protocol.112 In addition, gross pathology and histopathology study records were audited retrospectively by an independent QA contractor against study records submitted to the NTP Archives. Separate audits covered completeness and accuracy of the pathology data, pathology specimens, final pathology tables, and a draft of this NTP Toxicity Report. Audit procedures and findings are presented in the reports and are on file at the National Institute of Environmental Health Sciences. The audit findings were reviewed and assessed by DTT and NIOSH staff, and all comments were resolved or otherwise addressed during the preparation of this Toxicity Report.
Genetic Toxicology
The genetic toxicity of S. chartarum was assessed by determining the frequency of micronucleated erythrocytes in mouse peripheral blood. The protocol for these studies and the results are given in Appendix D.
The genetic toxicity studies have evolved from an earlier effort to develop a comprehensive database permitting a critical anticipation of a chemical’s carcinogenicity in experimental animals based on numerous considerations, including the relationship between the molecular structure of the chemical and its observed effects in short-term in vitro and in vivo genetic toxicity tests (structure-activity relationships). The short-term tests were developed originally to clarify proposed mechanisms of chemical-induced DNA damage, given the relationship between electrophilicity and mutagenicity,113 and the somatic mutation theory of cancer.114,115 Not all cancers, however, arise through genotoxic mechanisms.
Peripheral Blood Micronucleus Test
Micronuclei (literally “small nuclei” or Howell-Jolly bodies) are biomarkers of induced structural or numerical chromosomal alterations formed when acentric fragments or whole chromosomes fail to incorporate into either of two daughter nuclei during cell division.116,117 Acute in vivo bone marrow chromosome aberration and micronucleus tests appear to be less predictive of carcinogenicity than the Salmonella test.118,119 However, clearly positive results in long-term peripheral blood micronucleus tests have high predictivity for rodent carcinogenicity; a weak response in one sex only or negative results in both sexes in this assay do not correlate well with either negative or positive results in rodent carcinogenicity studies.120 Because of the theoretical and observed associations between induced genetic damage and adverse effects in somatic and germ cells, the determination of in vivo genetic effects is essential to the overall understanding of the risks associated with a particular exposure.
Results
Data Availability
All study data were evaluated. Data relevant for evaluating toxicological findings are presented here. All study data are available in the National Toxicology Program (NTP) Chemical Effects in Biological Systems (CEBS) database: https://doi.org/10.22427/NTP-DATA-TOX-107.121
Three-month Study
All female mice survived to the end of the study. One male mouse in the air control group suffered a hind limb wound and was euthanized on day 8 of exposure. All tissues defined in Table 1 were collected from all 10 males and 10 females following necropsy.
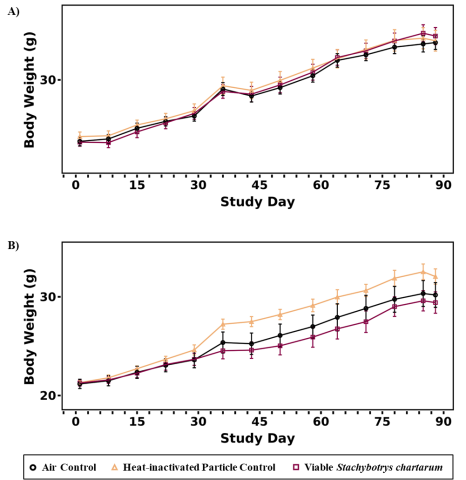
Final survival and body weight data are shown in Table 2 and Table 3. Terminal body weights of male and female mice exposed to viable S. chartarum conidia (spores) were not significantly different from the air control or the heat-inactivated particle control groups, nor were terminal body weights significantly different between the heat-inactivated particle control and air control groups. The body weight gain among groups of both male and female mice over the course of the study was similar, as illustrated in Figure 5. Clinical observations collected throughout the duration of the study revealed no remarkable findings (Appendix E).
A minimal but significant decrease (<1.3%) in mean cell volume was observed in the viable S. chartarum-exposed male mice compared to the air control and heat-inactivated particle control groups (Appendix E). This change was not observed in the viable S. chartarum-exposed female mice and was not considered exposure related. Eosinophil counts were significantly increased in the viable S. chartarum-exposed male mice relative to the air control and heat-inactivated particle control groups (Table 4). Although not significant, eosinophil counts were higher in the viable S. chartarum-exposed female mice relative to the air control and heat-inactivated particle control groups.
Absolute and relative lung weights were significantly increased in viable S. chartarum-exposed female mice compared to the air control group (33% and 35%, respectively) (Table 5). Although not significant, the absolute and relative lung weights of male mice exposed to viable S. chartarum were also higher compared to the air control mice (11% and 5%, respectively) (Table 5).
Histopathology
This section describes the statistically significant or biologically noteworthy changes in the incidences of nonneoplastic lesions of the larynx, lung, and bronchial lymph node.
Gross pathology was evaluated for the organs listed in Table 1. There were no gross lesions identified in any organs evaluated in the male and female groups (Appendix E).
Nonneoplastic lesions in the larynx, lung, and bronchial lymph nodes of viable S. chartarum-exposed male and female mice were present in increased numbers compared to air and heat-inactivated particle control groups (Table 6, Table 7, Table 8). The incidences of nonneoplastic lesions were also higher in the larynx of heat-inactivated particle control-exposed female mice compared to the air control.
The presence of fungal material in tissue sections was not observed following hematoxylin and eosin staining. Histological sections of aorta, gallbladder, heart, liver, lung, bronchial lymph node, and spleen were stained with Grocott’s methenamine silver (GMS) stain to detect fungal elements. No fungal material was observed in the aorta, gallbladder, heart, liver, or spleen sections.
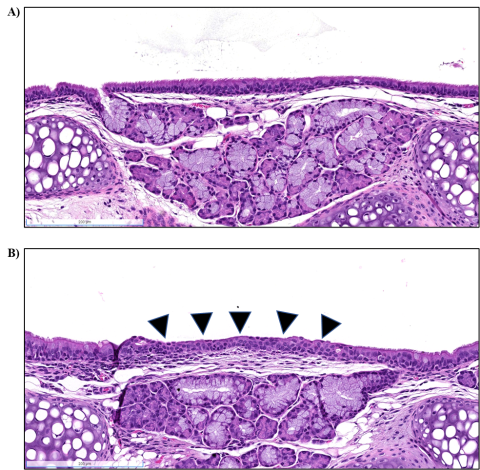
Larynx: In the larynx of viable S. chartarum-exposed male and female mice, the incidence of squamous metaplasia of the epiglottis was significantly increased compared to the air control (males and females) or heat-inactivated particle control (males) groups (Table 6). Squamous metaplasia of the epiglottis was observed in 3/10 female mice in the heat-inactivated particle control group. The lesions were present at the base of the epiglottis and were characterized by the replacement of normal ciliated cuboidal to columnar epithelium with nonkeratinized squamous epithelium (Figure 6). The metaplasia was graded as minimal (severity score of 1) in all instances and involved <50% of the epithelium overlying the glands at the base of the epiglottis.
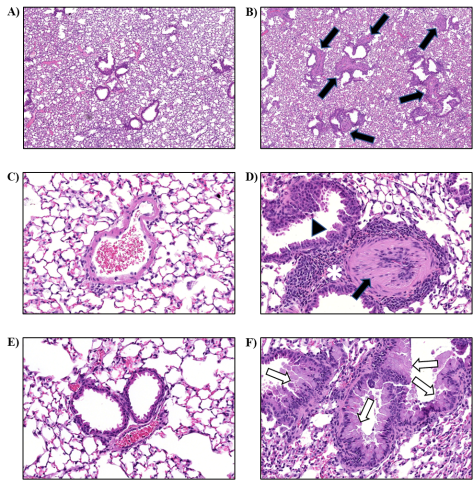
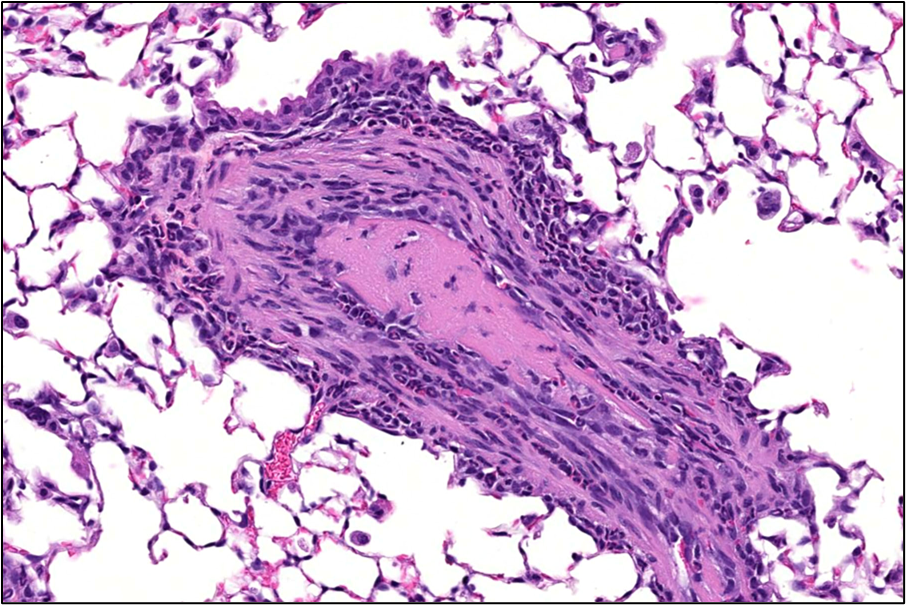
Lung: Numerous changes were observed in the lungs of viable S. chartarum-exposed mice compared to the air control and heat-inactivated particle control groups, which both displayed no lesions. Perivascular chronic-active inflammation was evident in all the viable S. chartarum-exposed males and females and was not observed in the air control or heat-inactivated particle control groups (Table 7). Chronic-active inflammation was characterized by the predominantly perivascular accumulations of lymphocytes, neutrophils, eosinophils, and macrophages in pulmonary artery walls, with extension into the immediately surrounding interstitial tissue (Figure 7).
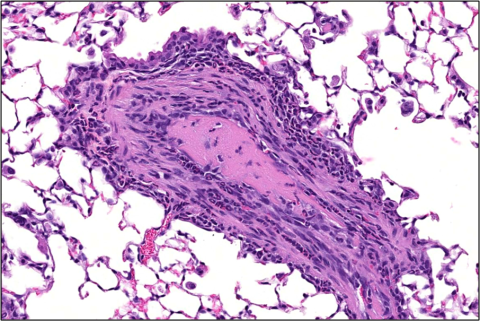
The perivascular chronic-active inflammation in viable S. chartarum-exposed mice was associated with thickening of the small- and medium-sized pulmonary arteries (medial hypertrophy, Figure 7). Mild to moderate medial hypertrophy of pulmonary arteries was present in all male and female viable S. chartarum-exposed mice and was not observed in the air control or heat-inactivated particle control groups (Table 6). The medial hypertrophy was characterized by circumferential thickening of the tunica media by concentric layers of hypertrophied and hyperplastic smooth muscle cells of the affected artery walls, accompanied by narrowing of the lumens. In three viable S. chartarum-exposed males and one viable S. chartarum-exposed female (Table 6), the artery medial hypertrophy was associated with very low numbers of intravascular thrombi (Figure 8). Medial hypertrophy and perivascular chronic-active inflammation severity was graded based upon the proportion of pulmonary arteries affected as follows: minimal severity (1) was <10% affected, mild (2) was 10%–40% affected, moderate (3) was 41%–75% affected, and marked (4) was >75% affected.
Bronchiolar epithelial hyperplasia was observed in all viable S. chartarum-exposed male and female mice and was not observed in the air control or heat-inactivated particle control groups (Table 7). Minimal bronchiolar epithelial hyperplasia was characterized by crowded, plump, often piled (2–5 cell layers thick), cuboidal, ciliated epithelial cells lining terminal bronchioles, with extension into adjacent alveolar septa (Figure 7). In most cases, the bronchiolar epithelial hyperplasia involved <10% of bronchioles and was considered minimal (1) but was graded as mild (2) in one male viable S. chartarum-exposed mouse in which 10%–20% of the bronchioles were affected.
Lymphocyte hyperplasia of the bronchus-associated lymphoid tissue (BALT) in the lung was observed in 9/10 viable S. chartarum-exposed males and females and was not observed in the air control or heat-inactivated particle control groups (Table 7). Lymphocyte hyperplasia of the BALT consisted of an increase in the size and/or number of aggregates of lymphocytes adjacent to bronchi when compared with that observed in air control animals. Air control animals had 0 to 1 lymphoid aggregates (most had 0). The severity of BALT lymphocyte hyperplasia was minimal (1) in all cases.
Histiocytic cellular infiltration in the lung was observed in all viable S. chartarum-exposed male and female animals (Table 7). The infiltration was characterized by a minimal to moderate increase in the numbers of diffusely scattered macrophages and rare multinucleated giant cells within alveolar spaces. The lesion severity was graded based upon the proportion of lung affected as follows: minimal severity (1) had <10% of the lung affected, mild (2) had 10%–40% affected, moderate (3) had 41%–75% affected, and marked (4) had >75% affected.
Goblet cell metaplasia was observed in all viable S. chartarum-exposed male and female mice and was not observed in the air control or heat-inactivated particle control groups (Table 7). Goblet cell metaplasia affected most to all of the epithelium in an individual airway and was characterized by increased numbers of goblet cells in the epithelium lining the bronchi and/or bronchioles (Figure 7). The severity of this lesion ranged from minimal to moderate and was graded based upon the proportion of airways affected as follows: minimal (1) had <10% of airways affected, mild (2) had 10%–40% affected, moderate (3) had 41%–75% affected, and marked (4) had >75% affected.
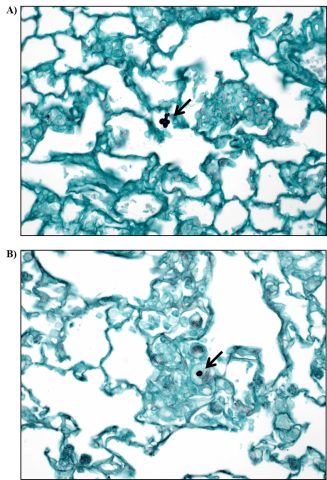
The lung sections of 9/10 male and 10/10 female viable S. chartarum-exposed mice stained positive for fungal material (Table 7). In the heat-inactivated particle control groups, GMS-positive fungal material was also observed in 1/10 male mice and 9/10 female mice (Table 7). GMS-positive structures consistent with viable S. chartarum conidia were observed in very low numbers (usually <5 throughout the entire lung section) and were round to oval, measuring 3 to 6 µm, and observed intracellularly within alveolar macrophages and occasionally free floating within the alveolar spaces (Figure 9).
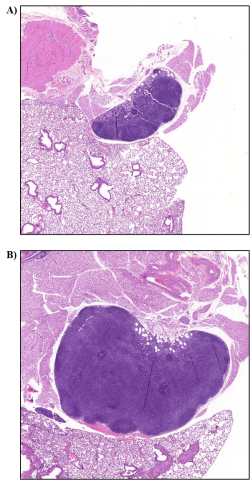
Bronchial lymph node: Bronchial, mandibular, mediastinal, and mesenteric lymph nodes were analyzed in all study mice. Histopathological analysis showed no visible lesions in the mandibular, mediastinal, or mesenteric lymph nodes. In the bronchial lymph node, lymphocyte hyperplasia was observed in two female viable S. chartarum-exposed mice but not in the viable S. chartarum-exposed male mice (Table 8). No lesions were observed in the bronchial lymph nodes of air control or heat-inactivated particle control mice. Lymphocyte hyperplasia was minimal in severity and characterized by increased numbers of lymphocytes within the cortex, paracortex, and medullary regions (Figure 10).
Very low numbers of conidia were also observed in the bronchial lymph node of the two female viable S. chartarum-exposed mice that also had lymphocyte hyperplasia of the bronchial lymph node (Table 8).
Genetic Toxicology
No significant increases in the frequencies of micronucleated reticulocytes and mature erythrocytes were observed in the peripheral blood of male or female B6C3F1/N mice exposed to either viable S. chartarum or heat-inactivated particle control conidia relative to the air control mice (Table D-1). Similarly, no significant increase in frequency was observed in mice exposed to viable S. chartarum conidia compared to mice exposed to the heat-inactivated particle control. In addition, no significant alteration in the percentage of reticulocytes was observed in these mice, suggesting no effects on hematopoiesis. These negative results in the micronucleus test indicate an absence of exposure-induced chromosomal damage in progenitor erythrocytes in the bone marrow of mice, along with an absence of generalized bone marrow toxicity. Data from all NTP genetic toxicity tests with S. chartarum are available in the NTP CEBS database: https://doi.org/10.22427/NTP-DATA-TOX-107.121
Discussion
Stachybotrys chartarum is a hydrophilic fungus that requires a cellulose-rich substrate and a high moisture content for optimal growth.35-38 This species is often identified in water-damaged indoor environments and has a worldwide distribution.35-38,122 Following infant acute idiopathic pulmonary hemorrhage cases,14 as well as the association with “sick building syndrome” and damp building-related illness,44,46,59,62-66 the adverse health effects following S. chartarum exposure have become a concern for public health. S. chartarum phialides produce conidia (spores) within a slime droplet that protects the conidia over a long period and inhibits aerosolization. Once dried, the conidia, hyphae, and fragments may be aerosolized within the breathing zone following abiotic or biotic disturbances. While most S. chartarum conidia are too large to deposit deep within the murine lung, this is not the case with the small respirable-size fungal fragments present within this test article.
Although mycotoxicosis and pulmonary immunological endpoints have been previously examined,29,101 the systemic toxicity following subchronic inhalation of viable S. chartarum remains undercharacterized. To evaluate the toxicological responses to S. chartarum, groups of 10 male and 10 female B6C3F1/N mice inhaled viable S. chartarum conidia, heat-inactivated S. chartarum conidia (biological particle control), or high-efficiency particulate air [filter] (HEPA)-filtered air (control) twice a week for 3 months. An estimated pulmonary dose of 1 × 104 S. chartarum conidia (viable and heat-inactivated conidia) was chosen for this study, which was the highest aerosolized dose that reproducibly induced a lymphoproliferative response and was the same concentration and schedule used for a previously published pulmonary immunology study.27 This estimated lung burden would be equivalent to that of a worker breathing a constant workplace air concentration of 3.45 × 106 conidia/m3 (146 μg/m3) for 1 week, working 8 hours per day for 5 days or equivalent to a worker breathing 2.3 × 104 conidia/m3 (0.97 μg/m3) for 8 hours per day, 5 days per week over a 40-year working career. This concentration is comparable to the occupational exposure concentration of 103–107 conidia/m3, which has been reported to result in toxicosis.43 In addition, studies involving S. chartarum-contaminated residences have also reported spore concentrations around 4 × 103 conidia/m3 during remediation86 and 103 conidia/mg of dust collected from contaminated areas.123
In the current study, inhalation exposure to an estimated pulmonary dose of 1 × 104 viable S. chartarum conidia twice a week for 3 months did not elicit overt toxicity or result in >10% body weight loss compared to the air control group for male or female mice. Additionally, the terminal body weights of mice exposed to either viable S. chartarum or the heat-inactivated particle control were not significantly different from those of the air control group. Although not significant, the absolute and relative lung weights of male mice exposed to viable S. chartarum were slightly higher compared to the air control group, whereas the viable S. chartarum-exposed female mice had significantly increased absolute and relative lung weights compared to the air control group.
No gross lesions were present following exposure; however, the incidences of nonneoplastic lesions were significantly increased in mice exposed to viable S. chartarum. These histopathological changes were evident in the lung and, to a lesser extent, in the larynx and bronchial lymph nodes of viable S. chartarum-exposed mice. Within the rest of the respiratory tract, no histopathological changes were observed in the nose or trachea. In the larynx of both male and female viable S. chartarum-exposed mice, the incidence of squamous metaplasia at the base of the epiglottis was significantly increased compared to the air control mice. In addition, squamous metaplasia at the base of the epiglottis was also observed in the larynx of three female mice exposed to the heat-inactivated particle control. The squamous metaplasia of the larynx epithelium at the base of the epiglottis included replacement of ciliated cuboidal to columnar cells with nonkeratinized stratified squamous epithelium, which is in response to chronic irritation elicited from the viable S. chartarum exposure. The National Toxicology Program (NTP) study detailing the toxicological responses following a subchronic Aspergillus fumigatus exposure also reported similar laryngeal squamous metaplasia at the base of the epiglottis in mice exposed to viable A. fumigatus.30 Literature has suggested that the larynx is the site that often responds to repeated inhalation of substances, including fungi, and that the resultant squamous metaplasia is frequently of minimal severity.30,124 Additionally, in the bronchial lymph node, lymphocyte hyperplasia was observed in two viable S. chartarum-exposed female mice, but no lesions were observed in S. chartarum-exposed male mice or in the bronchial lymph nodes of air control or heat-inactivated particle control mice.
Histopathological analysis of the lungs of viable S. chartarum-exposed males and females showed nonneoplastic lesions, including perivascular chronic-active inflammation, histiocytic cellular infiltration, medial hypertrophy of pulmonary arteries, thrombi in arteries with medial hypertrophy, hyperplasia of bronchiolar epithelium, goblet cell metaplasia, and bronchus-associated lymphoid tissue (BALT) lymphocyte hyperplasia. The mild to moderate medial hypertrophy affected small- and medium-sized pulmonary arteries and was characterized by circumferential thickening of the tunica media and luminal narrowing and was accompanied by accumulations of inflammatory cells. Intravascular thrombi, which were present in three males and one female exposed to S chartarum, were observed in very low numbers of arterioles that were remodeled as a result of medial hypertrophy. Despite the low incidence and number of affected vessels, the thrombi were considered likely exposure related because they were associated with vascular changes of luminal narrowing and medial hypertrophy. Similar to the current study, previously reported S. chartarum exposure via inhalation (twice per week for 4 weeks or 3 months) in female B6C3F1/N mice resulted in peribronchiolar and perivascular inflammation, alveolar histiocytosis, bronchiolar epithelial cell hyperplasia, as well as pulmonary arterial remodeling characterized by the thickening of the pulmonary arterial wall and narrowing of the lumen.27 Neither the air control nor heat-inactivated particle control groups in this study, as well as the previously published pulmonary immunology exposure study,27 presented evidence of medial hypertrophy of arteries. In addition, separate studies showed the development of pulmonary arterial hypertension in mice following repeated intratracheal instillation of S. chartarum,25,125 accompanied by significantly increased right ventricular systolic pressure and right ventricular hypertrophy in exposed male ddY mice compared to the control mice.25 Although cardiac function and pathology were not evaluated in the current study, the heart weights of the S. chartarum-exposed mice were not significantly different compared to the air control group, which is in contrast to the study conducted by Ochiai et al.25 These divergent results could be explained by the difference in delivery methods or the difference in mouse or fungal strain. Interestingly, cessation of S. chartarum exposure via intratracheal instillation resulted in the resolution of the pulmonary remodeling125; however, further investigation is warranted to determine whether this resolution occurs following repeated inhalation of viable S. chartarum conidia.
In comparison to the current study, an NTP study conducted in collaboration with the National Institute for Occupational Safety and Health (NIOSH) describing the toxicological responses following a subchronic viable A. fumigatus exposure showed similar pulmonary arterial medial hypertrophy.30 In addition, chronic-active inflammation, defined as the increased presence of neutrophils and perivascular infiltrates, including alveolar macrophages and eosinophils, was observed in mice exposed to viable A. fumigatus,30 and similar cell populations were observed following subchronic viable S. chartarum exposure. The significantly increased eosinophils observed in male mice may correspond to the chronic-active inflammation in the lung following subchronic viable S. chartarum exposure.
Perivascular chronic-active inflammation observed in the current study was characterized by an influx of inflammatory cell populations, including lymphocytes, neutrophils, eosinophils, and macrophages in the pulmonary arterial walls with extension into the immediately adjacent interstitial tissue and alveoli. In addition, histiocytic cellular infiltrates were observed in all viable S. chartarum-exposed male and female mice. All findings were also observed in female B6C3F1/N mice in a previously published study, following a 3-month exposure of viable S. chartarum via inhalation.27 Croston and colleagues showed, in the analysis of the bronchoalveolar lavage fluid (BALF), a significant increase in neutrophils, macrophages, and eosinophils at 24 hours and 48 hours post final exposure following a 3-month exposure to S. chartarum.27 In agreement, Nagayoshi et al. showed a significant increase in eosinophils accompanied by high concentrations of T helper 2 cell (Th2)-associated cytokines, interleukin (IL)-4 and IL-5, in the BALF following a 12-week intratracheal instillation of S. chartarum in male ddY mice.125 Using male BALB/c mice, Rosenblum Lichtenstein and colleagues also showed significantly increased BALF neutrophils, lymphocytes, and eosinophils and significantly increased IL-4, IL-5, and IL-6, supporting a Th2 response following a 7-week S. chartarum intratracheal instillation.126 Using the same mouse strain, Leino et al. reported significantly increased inflammatory cells including monocytes, neutrophils, and lymphocytes in response to intranasal instillation (twice a week for 3 weeks) of 1 × 105 satratoxin-producing and nonsatratoxin-producing S. chartarum spores in female mice.127 No inflammatory cellular infiltrates were observed following exposure to 1 × 103 satratoxin-producing and nonsatratoxin-producing S. chartarum spores.127 Additionally, proinflammatory cytokines were induced in the lungs of S. chartarum-exposed mice,127 and in contrast to previously published data,125,126 there was no significant increase in the expression of hallmark Th1 or Th2 cytokines. Although cytokine expression was not measured in the current study, Croston and colleagues showed that a 4-week exposure to viable S. chartarum resulted in a Th2-mediated response that switched to a mixed T-cell response after 3 months of exposure, as evidenced by significantly increased ifng (interferon-gamma; IFN-γ), il2 (IL-2), and il13 (IL-13) gene expression in the lungs of exposed mice.27 This mixed T-cell response is in contrast to other literature reporting a Th2-mediated response following 7 weeks126 and 12 weeks125 of S. chartarum exposure and, as previously stated, could be due to the difference in exposure methods or the different mouse and fungal strains used for the studies. While literature describes the evidence of Th2-mediated pulmonary arterial remodeling after 12 weeks of S. chartarum exposure,25,27,125 this pathological observation has been observed as early as 4 weeks following exposure to the same exposure schedule and strain of S. chartarum used in the current study.27 Ochiai et al.25 reported the development of pulmonary arterial thickening in S. chartarum-exposed mice after 4 weeks of intratracheal instillation. Previous studies suggest that the duration of S. chartarum exposures,27,126 the route of delivery,23-26,128 fungal strain,27 and the animal model employed129 could influence the resulting immune responses and lung pathologies.
Studies have indicated that different strains of mice respond differently following S. chartarum exposure.129 Specifically, BALB/c (Th2-biased) mice showed a different lung injury profile and inflammatory-associated biochemical and cellular profile than C57BL/6J (Th1-biased)126,129 and C3H/HeJ mice.129 The hybrid B6C3F1/N mouse strain used in the current study has been widely used to study systemic toxicological responses and was selected for consistency with other NTP toxicology studies. This strain is resistant to disease and has low spontaneous disease rates compared with other strains and hybrids.130 Although the female parent of the hybrid B6C3F1/N mouse strain, C57BL/6, is Th1-biased, histopathological results of the airways are consistent with previously observed histopathology in other strains of mice, including ddY mice.25 Moreover, the ddY mouse strain has been previously used as a model of S. chartarum-induced idiopathic pulmonary arterial hypertension.131 For the studies using the ddY mouse model, the S. chartarum test article was administered via intratracheal instillation.25,125,131 Compared to intratracheal and intranasal instillation or liquid aerosol inhalation that deliver conidia in a liquid suspension,23-26,128 which can potentially modify the viability and components of the conidia,42 the current study used a previously developed system to deliver dry aerosolized conidia to mice housed in nose-only exposure chambers, simulating a natural exposure that could be experienced in a S. chartarum-contaminated environment. Although the S. chartarum test article in the current study was not grown on building materials as has been previously used, the use of rice as a nutrient-rich growth substrate eliminates the potential interference of building material exposure to the mice housed in the exposure units. These differences in the administration of the S. chartarum conidia could partly account for the contrasting results observed among the different exposure studies. In addition, the strain of S. chartarum used in the study may also be a parameter that could influence the pulmonary responses following repeated exposure. As previously reported,27 a higher-fragmenting strain of S. chartarum (IBT 9460) elicited an earlier pulmonary immune response and pulmonary arterial remodeling compared to a lesser fragmenting strain of S. chartarum (IBT 7711), indicating that smaller fragments may deposit deeper in the lungs and induce these responses. Therefore, the higher-fragmenting S. chartarum strain (IBT 9460) was selected for toxicological evaluation in the current study. Furthermore, the cultivation and aerosolization of IBT 9460 has been previously optimized.28 In the study conducted by Lemons et al., both the viable S. chartarum test article (IBT 9460) and the heat-inactivated particle control contained similar levels of verrucarol, a hydrolysis product of macrocyclic trichothecene, which were four times higher than the S. chartarum (IBT 7711) strain. In addition, the endotoxin level of the viable S. chartarum test article was near or below the limit of detection.28
In the current 3-month study, very low numbers of conidia were observed free in the alveoli or within alveolar macrophages of most viable S. chartarum-exposed mice (Figure 9) and in the bronchial lymph node of two viable S. chartarum-exposed female mice. Following a 4-week intratracheal instillation of S. chartarum spores in male ddY mice, spores were shown to be present for only 24 hours before being cleared by alveolar macrophages.125 In addition, fungal components were not visualized in left lung lobes following a 3-month inhalation exposure to S. chartarum in female B6C3F1 mice,27 which is contrary to the histopathological observations of the current study. Similar to the current study, A. fumigatus spores were detected within the lungs of mice following exposure; however, in a few cases, swollen conidia and germ tubes were detected, indicating active germination of the conidia,30,100,102 which was not observed in the current study.
Hyphal fragments can also be aerosolized within an indoor environment following abiotic or biotic disturbance. Studies have demonstrated that fungal fragments contribute to the fungal biomass within indoor environments.10,11 Studies of S. chartarum have predicted that fragments can be aerosolized at particle concentrations 500 times higher than spores.8 Furthermore, deposition of submicron fungal fragments deep within the human respiratory tract is 230–250 fold higher than that of spores, which would primarily be deposited in the upper respiratory tract.8 Croston and colleagues have recently proposed fragments as the primary driver of pulmonary immune responses following the inhalation of S. chartarum.27 Moreover, literature has suggested that the macrocyclic trichothecenes detected on S. chartarum fragments may contribute to the toxigenic health effects following exposure.37,47 In addition to macrocyclic trichothecenes, other biologically active products of S. chartarum, such as hemolysins, proteinases, glucans, and volatile organic compounds, could act in concert to contribute to the toxigenic health effects following exposure.37 A study conducted by Hudson et al. demonstrated pulmonary immune response differences in the magnitude and temporal interval following intratracheal instillation exposure to conidia of macrocyclic-trichothecene-producing (JS 58-17) and nonproducing chemotype (JS 58-06) S. chartarum.132 Additionally, Rand and colleagues determined that pulmonary tissue collected from CFW male mice following a single intratracheal instillation of S. chartarum spores responds differently compared to pure trichothecene exposure (isosatratoxin F), including significantly decreased alveolar air space and significantly increased erythrocytes in the intra-alveolar air space.133 Although mycotoxin profiles and concentrations were not evaluated in the current study, Lemons et al. showed similar levels of verrucarol in the viable and heat-inactivated particle control aerosols of the same strain of S. chartarum.28 Furthermore, Croston et al. showed that a more rapid immune response and pulmonary remodeling occurred following exposure to this higher-fragmenting and mycotoxin-producing strain of S. chartarum (IBT 9460).27 As the peer-reviewed literature has suggested, fragment size, mycotoxin content, or a combination of these factors could be responsible for the immune response and arterial remodeling observed; therefore, further research is warranted.134
Findings from the current 3-month inhalation study in B6C3F1/N mice included significantly increased absolute and relative lung weights in viable S. chartarum-exposed female mice, relative to the heat-inactivated control and air control groups accompanied by significantly increased incidences of nonneoplastic lesions. In males and females exposed to viable S. chartarum, nonneoplastic lesions were restricted mainly to the larynx and lung; however, there was also a low incidence of lymphocyte hyperplasia in the bronchial lymph nodes of viable S. chartarum-exposed female mice. In the larynx, nonneoplastic lesions consisted of squamous metaplasia of the epithelium localized at the base of the epiglottis. Lung histopathology findings were composed of several lesions, including medial hypertrophy of the pulmonary arteries/arterioles in viable S. chartarum-exposed mice but not in heat-inactivated particle control mice. These results correspond with the results from a previously published subchronic S. chartarum pulmonary immunology study conducted by NIOSH with the same exposure system that showed Th2-mediated pulmonary inflammation and pulmonary arterial remodeling after subchronic S. chartarum inhalation exposure.27 The results of the current study were also similar to the previously published NTP report detailing the toxicological responses to A. fumigatus exposure in which the incidence of nonneoplastic lesions was significantly increased in the larynx, lung, and bronchial lymph nodes of viable A. fumigatus-exposed mice.30 Specifically, the incidences of bronchial lymphocytic hyperplasia and plasma cell hyperplasia were significantly increased in viable A. fumigatus-exposed mice but not in viable S. chartarum-exposed mice, relative to either the air control or the heat-inactivated particle control groups.30 In both the current study of S. chartarum and the previously published NTP report of A. fumigatus, lungs of mice exposed to viable conidia displayed inflammation, medial hypertrophy of pulmonary arterioles and arteries, bronchiolar epithelium hyperplasia, goblet cell metaplasia, BALT lymphocyte hyperplasia, and the presence of very low numbers of conidia.
Under the conditions of this 3-month study, target organs identified in B6C3F1/N mice following inhalation exposure to viable S. chartarum conidia were the larynx, lung, and (to a lesser extent) the bronchial lymph nodes. The histopathological changes observed following viable S. chartarum exposure were not seen in the heat-inactivated particle control or air control mice. These results build on initial pulmonary immunology studies conducted at NIOSH using the same exposure parameters and demonstrate that the immunological responses and histopathology could be mediated by the viable S. chartarum bioaerosol.
References
1. Andersen B, Nielsen KF, Jarvis BB. Characterization of Stachybotrys from water-damaged buildings based on morphology, growth, and metabolite production. Mycologia. 2002; 94(3):392-403. DOI: 10.1080/15572536.2003.11833204 PubMed: 21156510
2. Kung’u J. Is black mold (Stachybotrys) a new emerging opportunistic human fungal pathogen? Mississauga, Ontario, Canada: Mold & Bacteria Consulting Laboratories; 2020. https://www.moldbacteria.com/mold/is-black-mold-stachybotrys-a-new-emerging-opportunistic-human-fungal-pathogen.html
3. Blackwell M. The fungi: 1, 2, 3 … 5.1 million species? Am J Bot. 2011; 98(3):426-438. DOI: 10.3732/ajb.1000298 PubMed: 21613136
4. Green BJ. Occupational fungal exposure in the United States. In: Viegas C, Viegas S, Gomes A, Täubel M, Sabino R, editors. Exposure to Microbiological Agents in Indoor and Occupational Environments. Cham, Switzerland: Springer; 2017. p. 3-33. DOI: 10.1007/978-3-319-61688-9_1
5. Gow NAR, Latge JP, Munro CA. The fungal cell wall: Structure, biosynthesis, and function. Microbiol Spectr. 2017; 5(3). DOI: 10.1128/microbiolspec.FUNK-0035-2016 PubMed: 28513415
6. Eduard W. Fungal spores: A critical review of the toxicological and epidemiological evidence as a basis for occupational exposure limit setting. Crit Rev Toxicol. 2009; 39(10):799-864. DOI: 10.3109/10408440903307333 PubMed: 19863384
7. Green BJ, Sercombe JK, Tovey ER. Fungal fragments and undocumented conidia function as new aeroallergen sources. J Allergy Clin Immunol. 2005; 115(5):1043-1048. DOI: 10.1016/j.jaci.2005.02.009 PubMed: 15867864
8. Cho SH, Seo SC, Schmechel D, Grinshpun SA, Reponen T. Aerodynamic characteristics and respiratory deposition of fungal fragments. Atmos Environ. 2005; 39(30):5454-5465. DOI: 10.1016/j.atmosenv.2005.05.042
9. Green BJ, Schmechel D, Summerbell RC. Aerosolized fungal fragments. In: Adan OCG, Samson RA, editors. Fundamentals of Mold Growth in Indoor Environments and Strategies for Healthy Living. Wageningen, Netherlands: Wageningen Academic Publishers; 2011. p. 211-243. DOI: 10.3920/9789086867226_009
10. Adhikari A, Reponen T, Rylander R. Airborne fungal cell fragments in homes in relation to total fungal biomass. Indoor Air. 2013; 23(2):142-147. DOI: 10.1111/j.1600-0668.2012.00799.x PubMed: 22804753
11. Reponen T, Seo SC, Grimsley F, Lee T, Crawford C, Grinshpun SA. Fungal fragments in moldy houses: A field study in homes in New Orleans and southern Ohio. Atmos Environ (1994). 2007; 41(37):8140-8149. DOI: 10.1016/j.atmosenv.2007.06.027 PubMed: 19050738
12. Afanou KA, Straumfors A, Skogstad A, Nilsen T, Synnes O, Skaar I, Hjeljord L, Tronsmo A, Green BJ, Eduard W. Submicronic fungal bioaerosols: High-resolution microscopic characterization and quantification. Appl Environ Microbiol. 2014; 80(22):7122-7130. DOI: 10.1128/AEM.01740-14 PubMed: 25217010
13. Gravesen S, Nielsen PA, Iversen R, Nielsen KF. Microfungal contamination of damp buildingss—examples of risk constructions and risk materials. Environ Health Perspect. 1999; 107 Suppl 3:505-508. DOI: 10.1289/ehp.99107s3505 PubMed: 10347000
14. Centers for Disease Control and Prevention (CDC). Acute pulmonary hemorrhage/hemosiderosis among infants — Cleveland, January 1993–November 1994. MMWR Morb Mortal Wkly Rep. 1994; 43(48):881-883. PubMed: 7969010
15. Centers for Disease Control and Prevention (CDC). Update: Pulmonary hemorrhage/hemosiderosis among infants — Cleveland, Ohio, 1993–1996. MMWR Morb Mortal Wkly Rep. 1997; 46(2):33-35. PubMed: 9011781
16. Institute of Medicine (IOM). Damp indoor spaces and health. Washington, DC: The National Academies Press; 2004. https://nap.nationalacademies.org/catalog/11011/damp-indoor-spaces-and-health
17. World Health Organization (WHO). WHO guidelines for indoor air quality: Dampness and mould. Geneva, Switzerland: World Health Organization, Regional Office for Europe; 2009. https://www.who.int/publications/i/item/9789289041683
18. Mendell MJ, Mirer AG, Cheung K, Tong M, Douwes J. Respiratory and allergic health effects of dampness, mold, and dampness-related agents: A review of the epidemiologic evidence. Environ Health Perspect. 2011; 119(6):748-756. DOI: 10.1289/ehp.1002410 PubMed: 21269928
19. Baxi SN, Portnoy JM, Larenas-Linnemann D, Phipatanakul W, Environmental Allergens Workgroup. Exposure and health effects of fungi on humans. J Allergy Clin Immunol Pract. 2016; 4(3):396-404. DOI: 10.1016/j.jaip.2016.01.008 PubMed: 26947460
20. Benedict K, Jackson BR, Chiller T, Beer KD. Estimation of direct healthcare costs of fungal diseases in the United States. Clin Infect Dis. 2019; 68(11):1791-1797. DOI: 10.1093/cid/ciy776 PubMed: 30204844
21. Nikulin M, Reijula K, Jarvis BB, Hintikka EL. Experimental lung mycotoxicosis in mice induced by Stachybotrys atra. Int J Exp Pathol. 1996; 77(5):213-218. DOI: 10.1046/j.1365-2613.1996.9250323.x PubMed: 8977373
22. Rao CY, Brain JD, Burge HA. Reduction of pulmonary toxicity of Stachybotrys chartarum spores by methanol extraction of mycotoxins. Appl Environ Microbiol. 2000; 66(7):2817-2821. DOI: 10.1128/AEM.66.7.2817-2821.2000 PubMed: 10877773
23. Rand TG, Mahoney M, White K, Oulton M. Microanatomical changes in alveolar type II cells in juvenile mice intratracheally exposed to Stachybotrys chartarum spores and toxin. Toxicol Sci. 2002; 65(2):239-245. DOI: 10.1093/toxsci/65.2.239 PubMed: 11812928
24. Yike I, Rand TG, Dearborn DG. Acute inflammatory responses to Stachybotrys chartarum in the lungs of infant rats: Time course and possible mechanisms. Toxicol Sci. 2005; 84(2):408-417. DOI: 10.1093/toxsci/kfi080 PubMed: 15647601
25. Ochiai E, Kamei K, Watanabe A, Nagayoshi M, Tada Y, Nagaoka T, Sato K, Sato A, Shibuya K. Inhalation of Stachybotrys chartarum causes pulmonary arterial hypertension in mice. Int J Exp Pathol. 2008; 89(3):201-208. DOI: 10.1111/j.1365-2613.2008.00585.x PubMed: 18460072
26. Korpi A, Kasanen JP, Raunio P, Kosma VM, Virtanen T, Pasanen AL. Effects of aerosols from nontoxic Stachybotrys chartarum on murine airways. Inhal Toxicol. 2002; 14(5):521-540. DOI: 10.1080/089583701753678607 PubMed: 12028806
27. Croston TL, Lemons AR, Barnes MA, Goldsmith WT, Orandle MS, Nayak AP, Germolec DR, Green BJ, Beezhold DH. Inhalation of Stachybotrys chartarum fragments induces pulmonary arterial remodeling. Am J Respir Cell Mol Biol. 2020; 62(5):563-576. DOI: 10.1165/rcmb.2019-0221OC PubMed: 31671270
28. Lemons AR, Croston TL, Goldsmith WT, Barnes MA, Jaderson MA, Park JH, McKinney W, Beezhold DH, Green BJ. Cultivation and aerosolization of Stachybotrys chartarum for modeling pulmonary inhalation exposure. Inhal Toxicol. 2019; 31(13-14):446-456. DOI: 10.1080/08958378.2019.1705939 PubMed: 31874574
29. Nayak AP, Croston TL, Lemons AR, Goldsmith WT, Marshall NB, Kashon ML, Germolec DR, Beezhold DH, Green BJ. Aspergillus fumigatus viability drives allergic responses to inhaled conidia. Ann Allergy Asthma Immunol. 2018; 121(2):200-210.e2. DOI: 10.1016/j.anai.2018.04.008 PubMed: 29660515
30. National Toxicology Program (NTP). NTP technical report on the toxicity studies of Aspergillus fumigatus administered by inhalation to B6C3F1/N mice (revised). Research Triangle Park, NC: U.S. Department of Health and Human Services, Public Health Service, National Toxicology Program; 2021. NTP Toxicity Report 100. DOI: 10.22427/NTP-TOX-100
31. Kirk PM, Cannon PF, Minter DW, Stalpers JA. Dictionary of the fungi. 10th ed. Wallingford, UK: CABI International; 2008.
32. Ehrenberg CG. Sylvae mycologicae Berolinensis [dissertation]. University of Berlin; 1818. https://www.google.com/books/edition/_/gO5hAAAAcAAJ?gbpv=1
33. Corda ACJ. Icones fungorum hucusque cognitorum. Auctore A. C. J. Corda. Pragæ, 1837. Ann Mag Nat Hist Ser 1. 1838; 2(7):61-63. DOI: 10.1080/00222933809496655
34. Izabel TdSS, da Cruz ACR, Barbosa FR, Ferreira SML, Marques MFO, Gusmão LFP. The genus Stachybotrys (anamorphic fungi) in the semi-arid region of Brazil. Rev Bras Bot. 2010; 33(3):479-487. DOI: 10.1590/S0100-84042010000300010
35. Andersson MA, Nikulin M, Köljalg U, Andersson MC, Rainey F, Reijula K, Hintikka EL, Salkinoja-Salonen M. Bacteria, molds, and toxins in water-damaged building materials. Appl Environ Microbiol. 1997; 63(2):387-393. DOI: 10.1128/aem.63.2.387-393.1997 PubMed: 9023919
36. Boutin-Forzano S, Charpin-Kadouch C, Chabbi S, Bennedjai N, Dumon H, Charpin D. Wall relative humidity: A simple and reliable index for predicting Stachybotrys chartarum infestation in dwellings. Indoor Air. 2004; 14(3):196-199. DOI: 10.1111/j.1600-0668.2004.00233.x PubMed: 15104787
37. Pestka JJ, Yike I, Dearborn DG, Ward MDW, Harkema JR. Stachybotrys chartarum, trichothecene mycotoxins, and damp building-related illness: New insights into a public health enigma. Toxicol Sci. 2008; 104(1):4-26. DOI: 10.1093/toxsci/kfm284 PubMed: 18007011
38. Grant C, Hunter CA, Flannigan B, Bravery AF. The moisture requirements of moulds isolated from domestic dwellings. Int Biodeterior. 1989; 25(4):259-284. DOI: 10.1016/0265-3036(89)90002-X
39. Frazer S, Magan N, Aldred D. The influence of water activity and temperature on germination, growth and sporulation of Stachybotrys chartarum strains. Mycopathologia. 2011; 172(1):17-23. DOI: 10.1007/s11046-011-9394-x PubMed: 21347692
40. Mahmoudi M, Gershwin ME. Sick building syndrome. III. Stachybotrys chartarum. J Asthma. 2000; 37(2):191-198. DOI: 10.3109/02770900009055442 PubMed: 10805208
41. Kuhn DM, Ghannoum MA. Indoor mold, toxigenic fungi, and Stachybotrys chartarum: Infectious disease perspective. Clin Microbiol Rev. 2003; 16(1):144-172. DOI: 10.1128/CMR.16.1.144-172.2003 PubMed: 12525430
42. Madsen AM, Larsen ST, Koponen IK, Kling KI, Barooni A, Karottki DG, Tendal K, Wolkoff P. Generation and characterization of indoor fungal aerosols for inhalation studies. Appl Environ Microbiol. 2016; 82(8):2479-2493. DOI: 10.1128/AEM.04063-15 PubMed: 26921421
43. Miller JD, Rand TG, Jarvis BB. Stachybotrys chartarum: Cause of human disease or media darling? Med Mycol. 2003; 41(4):271-291. DOI: 10.1080/1369378031000137350 PubMed: 12964721
44. Hossain MA, Ahmed MS, Ghannoum MA. Attributes of Stachybotrys chartarum and its association with human disease. J Allergy Clin Immunol. 2004; 113(2):200-208. DOI: 10.1016/j.jaci.2003.12.018 PubMed: 14767429
45. Hodgson M, Dearborn DG. Human pulmonary disease and exposure to Stachybotrys chartarum and other toxigenic fungi. J Occup Environ Med. 2002; 44(8):705-707. DOI: 10.1097/00043764-200208000-00004 PubMed: 12185790
46. Jarvis BB, Sorenson WG, Hintikka EL, Nikulin M, Zhou Y, Jiang J, Wang S, Hinkley S, Etzel RA, Dearborn D. Study of toxin production by isolates of Stachybotrys chartarum and Memnoniella echinata isolated during a study of pulmonary hemosiderosis in infants. Appl Environ Microbiol. 1998; 64(10):3620-3625. DOI: 10.1128/AEM.64.10.3620-3625.1998 PubMed: 9758776
47. Brasel TL, Martin JM, Carriker CG, Wilson SC, Straus DC. Detection of airborne Stachybotrys chartarum macrocyclic trichothecene mycotoxins in the indoor environment. Appl Environ Microbiol. 2005; 71(11):7376-7388. DOI: 10.1128/AEM.71.11.7376-7388.2005 PubMed: 16269780
48. Fog Nielsen K. Mycotoxin production by indoor molds. Fungal Genet Biol. 2003; 39(2):103-117. DOI: 10.1016/S1087-1845(03)00026-4 PubMed: 12781669
49. Andersen B, Nielsen KF, Thrane U, Szaro T, Taylor JW, Jarvis BB. Molecular and phenotypic descriptions of Stachybotrys chlorohalonata sp. nov. and two chemotypes of Stachybotrys chartarum found in water-damaged buildings. Mycologia. 2003; 95(6):1227-1238. DOI: 10.1080/15572536.2004.11833031 PubMed: 21149024
50. Jarvis BB, Lee YW, Cömezoglu SN, Yatawara CS. Trichothecenes produced by Stachybotrys atra from eastern Europe. Appl Environ Microbiol. 1986; 51(5):915-918. DOI: 10.1128/aem.51.5.915-918.1986 PubMed: 3729393
51. Gao P, Martin J. Volatile metabolites produced by three strains of Stachybotrys chartarum cultivated on rice and gypsum board. Appl Occup Environ Hyg. 2002; 17(6):430-436. DOI: 10.1080/10473220290035462 PubMed: 12049433
52. Forgacs J. Stachybotryotoxicosis. In: Kadis S, Ciegler A, Ajl SJ, editors. Microbial Toxins: A Comprehensive Treatise, Volume VIII: Fungal Toxins. New York, NY: Academic Press; 1972. p. 95-128.
53. Drobotko VG. Stachybotryotoxicosis: A new disease of horses and humans. Am Rev Sov Med. 1945; 2(3):238-242.
54. Akkmeteli MA. Epidemiological features of the mycotoxicoses. Ann Nutr Aliment. 1977; 31(4-6):957-975. PubMed: 77650
55. Croft WA, Jarvis BB, Yatawara CS. Airborne outbreak of trichothecene toxicosis. Atmos Environ. 1986; 20(3):549-552. DOI: 10.1016/0004-6981(86)90096-X
56. Hodgson MJ, Morey P, Leung WY, Morrow L, Miller D, Jarvis BB, Robbins H, Halsey JF, Storey E. Building-associated pulmonary disease from exposure to Stachybotrys chartarum and Aspergillus versicolor. J Occup Environ Med. 1998; 40(3):241-249. DOI: 10.1097/00043764-199803000-00006 PubMed: 9531095
57. Johanning E. Health problems related to fungal exposure: The example of toxigenic Stachybotrys chartarum (atra). In: Johanning E, Yang CS, editors. Fungi and Bacteria in Indoor Air Environments: Health Effects, Detection, and Remediation: Proceedings of the International Conference, Saratoga Springs, New York, October 6-7, 1994. Latham, NY: Eastern New York Occupational Health Program; 1995. p. 201-208.
58. Johanning E, Biagini R, Hull D, Morey P, Jarvis B, Landsbergis P. Health and immunology study following exposure to toxigenic fungi (Stachybotrys chartarum) in a water-damaged office environment. Int Arch Occup Environ Health. 1996; 68(4):207-218. DOI: 10.1007/s004200050052 PubMed: 8738349
59. Cooley JD, Wong WC, Jumper CA, Straus DC. Correlation between the prevalence of certain fungi and sick building syndrome. Occup Environ Med. 1998; 55(9):579-584. DOI: 10.1136/oem.55.9.579 PubMed: 9861178
60. Etzel R, Rylander R. Indoor mold and children’s health. Environ Health Perspect. 1999; 107 Suppl 3:463. DOI: 10.1289/ehp.107-1566224 PubMed: 10346994
61. Tuomi T, Reijula K, Johnsson T, Hemminki K, Hintikka EL, Lindroos O, Kalso S, Koukila-Kähkölä P, Mussalo-Rauhamaa H, Haahtela T. Mycotoxins in crude building materials from water-damaged buildings. Appl Environ Microbiol. 2000; 66(5):1899-1904. DOI: 10.1128/AEM.66.5.1899-1904.2000 PubMed: 10788357
62. Li DW, Yang CS. Fungal contamination as a major contributor to sick building syndrome. Adv Appl Microbiol. 2004; 55:31-112. DOI: 10.1016/S0065-2164(04)55002-5 PubMed: 15350790
63. Hintikka EL. The role of stachybotrys in the phenomenon known as sick building syndrome. Adv Appl Microbiol. 2004; 55:155-173. DOI: 10.1016/S0065-2164(04)55005-0 PubMed: 15350793
64. Dearborn DG, Yike I, Sorenson WG, Miller MJ, Etzel RA. Overview of investigations into pulmonary hemorrhage among infants in Cleveland, Ohio. Environ Health Perspect. 1999; 107 Suppl 3:495-499. DOI: 10.1289/ehp.99107s3495 PubMed: 10346998
65. Dearborn DG, Smith PG, Dahms BB, Allan TM, Sorenson WG, Montana E, Etzel RA. Clinical profile of 30 infants with acute pulmonary hemorrhage in Cleveland. Pediatrics. 2002; 110(3):627-637. DOI: 10.1542/peds.110.3.627 PubMed: 12205270
66. Fung F, Clark R, Williams S. Stachybotrys, a mycotoxin-producing fungus of increasing toxicologic importance. J Toxicol Clin Toxicol. 1998; 36(1-2):79-86. DOI: 10.3109/15563659809162592 PubMed: 9541050
67. Johanning E, Morey PR, Jarvis BB. Clinical-epidemiological investigation of health effects caused by Stachybotrys atra building contamination. In: Jaakkola JJK, Ilmarinen R, Seppanen O, editors. Indoor Air '93: Proceedings of the 6th International Conference on Indoor Air Quality and Climate, Volume 1: Health effects. Helsinki, Finland: Indoor Air '93; 1993. p. 225-230.
68. Etzel RA, Montaña E, Sorenson WG, Kullman GJ, Allan TM, Dearborn DG. Acute pulmonary hemorrhage in infants associated with exposure to Stachybotrys atra and other fungi. Arch Pediatr Adolesc Med. 1998; 152(8):757-762. DOI: 10.1001/archpedi.152.8.757 PubMed: 9701134
69. Kozak PP Jr, Gallup J, Cummins LH, Gillman SA. Currently available methods for home mold surveys. II. Examples of problem homes surveyed. Ann Allergy. 1980; 45(3):167-176. PubMed: 6998332
70. Ochiai E, Kamei K, Hiroshima K, Watanabe A, Hashimoto Y, Sato A, Ando A. The pathogenicity of Stachybotrys chartarum. Nihon Ishinkin Gakkai Zasshi. 2005; 46(2):109-117. DOI: 10.3314/jjmm.46.109 PubMed: 15864257
71. Semis M, Dadwal SS, Tegtmeier BR, Wilczynski SP, Ito JI, Kalkum M. First case of invasive Stachybotrys sinusitis. Clin Infect Dis. 2021; 72(8):1386-1391. DOI: 10.1093/cid/ciaa231 PubMed: 32155243
72. Nelson BD. APSnet feature articles: Stachybotrys chartarum: The toxic indoor mold. St Paul, MN: American Phytopathological Society; 2001. https://www.apsnet.org/edcenter/apsnetfeatures/Pages/Stachybotrys.aspx
73. Andrássy K, Horváth I, Lakos T, Töke Z. [Mass incidence of mycotoxicoses in Hajdu-Bihar County]. Mykosen. 1980; 23(3):130-133. DOI: 10.1111/j.1439-0507.1980.tb02593.x PubMed: 7191050
74. El-Maghraby OMO, Bean GA, Jarvis BB, Aboul-Nasr MB. Macrocyclic trichothecenes produced by Stachybotrys isolated from Egypt and eastern Europe. Mycopathologia. 1991; 113(2):109-115. DOI: 10.1007/BF00442419 PubMed: 2034259
75. Gregory L, Pestka JJ, Dearborn DG, Rand TG. Localization of satratoxin-G in Stachybotrys chartarum spores and spore-impacted mouse lung using immunocytochemistry. Toxicol Pathol. 2004; 32(1):26-34. DOI: 10.1080/01926230490260790 PubMed: 14713545
76. Brasel TL, Douglas DR, Wilson SC, Straus DC. Detection of airborne Stachybotrys chartarum macrocyclic trichothecene mycotoxins on particulates smaller than conidia. Appl Environ Microbiol. 2005; 71(1):114-122. DOI: 10.1128/AEM.71.1.114-122.2005 PubMed: 15640178
77. Górny RL, Reponen T, Willeke K, Schmechel D, Robine E, Boissier M, Grinshpun SA. Fungal fragments as indoor air biocontaminants. Appl Environ Microbiol. 2002; 68(7):3522-3531. DOI: 10.1128/AEM.68.7.3522-3531.2002 PubMed: 12089037
78. Aleksic B, Draghi M, Ritoux S, Bailly S, Lacroix M, Oswald IP, Bailly JD, Robine E. Aerosolization of mycotoxins after growth of toxinogenic fungi on wallpaper. Appl Environ Microbiol. 2017; 83(16):e01001-17. DOI: 10.1128/AEM.01001-17 PubMed: 28646113
79. Jarvis BB. Macrocyclic trichothecenes. In: Sharma RP, Salunkhe DK, editors. Mycotoxins and Phytoalexins. Boca Raton, FL: CRC Press; 1991. p. 361-421.
80. Sorenson WG, Frazer DG, Jarvis BB, Simpson J, Robinson VA. Trichothecene mycotoxins in aerosolized conidia of Stachybotrys atra. Appl Environ Microbiol. 1987; 53(6):1370-1375. DOI: 10.1128/aem.53.6.1370-1375.1987 PubMed: 3496850
81. Yang GH, Jarvis BB, Chung YJ, Pestka JJ. Apoptosis induction by the satratoxins and other trichothecene mycotoxins: Relationship to ERK, p38 MAPK, and SAPK/JNK activation. Toxicol Appl Pharmacol. 2000; 164(2):149-160. DOI: 10.1006/taap.1999.8888 PubMed: 10764628
82. Islam Z, Harkema JR, Pestka JJ. Satratoxin G from the black mold Stachybotrys chartarum evokes olfactory sensory neuron loss and inflammation in the murine nose and brain. Environ Health Perspect. 2006; 114(7):1099-1107. DOI: 10.1289/ehp.8854 PubMed: 16835065
83. Mason CD, Rand TG, Oulton M, MacDonald JM, Scott JE. Effects of Stachybotrys chartarum (atra) conidia and isolated toxin on lung surfactant production and homeostasis. Nat Toxins. 1998; 6(1):27-33. DOI: 10.1002/(SICI)1522-7189(199802)6:1<27::AID-NT6>3.0.CO;2-B PubMed: 9851509
84. Kankkunen P, Rintahaka J, Aalto A, Leino M, Majuri ML, Alenius H, Wolff H, Matikainen S. Trichothecene mycotoxins activate inflammatory response in human macrophages. J Immunol. 2009; 182(10):6418-6425. DOI: 10.4049/jimmunol.0803309 PubMed: 19414795
85. Vesper SJ, Dearborn DG, Yike I, Sorenson WG, Haugland RA. Hemolysis, toxicity, and randomly amplified polymorphic DNA analysis of Stachybotrys chartarum strains. Appl Environ Microbiol. 1999; 65(7):3175-3181. DOI: 10.1128/AEM.65.7.3175-3181.1999 PubMed: 10388719
86. Vesper S, Dearborn DG, Yike I, Allan T, Sobolewski J, Hinkley SF, Jarvis BB, Haugland RA. Evaluation of Stachybotrys chartarum in the house of an infant with pulmonary hemorrhage: Quantitative assessment before, during, and after remediation. J Urban Health. 2000; 77(1):68-85. DOI: 10.1007/BF02350963 PubMed: 10741843
87. Vesper SJ, Magnuson ML, Dearborn DG, Yike I, Haugland RA. Initial characterization of the hemolysin stachylysin from Stachybotrys chartarum. Infect Immun. 2001; 69(2):912-916. DOI: 10.1128/IAI.69.2.912-916.2001 PubMed: 11159985
88. Vesper SJ, Vesper MJ. Stachylysin may be a cause of hemorrhaging in humans exposed to Stachybotrys chartarum. Infect Immun. 2002; 70(4):2065-2069. DOI: 10.1128/IAI.70.4.2065-2069.2002 PubMed: 11895972
89. Elidemir O, Colasurdo GN, Rossmann SN, Fan LL. Isolation of Stachybotrys from the lung of a child with pulmonary hemosiderosis. Pediatrics. 1999; 104(4 Pt 1):964-966. DOI: 10.1542/peds.104.4.964 PubMed: 10506242
90. Rand TG, Flemming J, Miller JD, Womiloju TO. Comparison of inflammatory responses in mouse lungs exposed to atranones A and C from Stachybotrys chartarum. J Toxicol Environ Health A. 2006; 69(13):1239-1251. DOI: 10.1080/15287390500360307 PubMed: 16754538
91. Fung F, Hughson WG. Health effects of indoor fungal bioaerosol exposure. Appl Occup Environ Hyg. 2003; 18(7):535-544. DOI: 10.1080/10473220301451 PubMed: 12791550
92. Hardin BD, Kelman BJ, Saxon A. Adverse human health effects associated with molds in the indoor environment. J Occup Environ Med. 2003; 45(5):470-478. DOI: 10.1097/00043764-200305000-00006 PubMed: 12762072
93. Jagels A, Lindemann V, Ulrich S, Gottschalk C, Cramer B, Hübner F, Gareis M, Humpf HU. Exploring secondary metabolite profiles of Stachybotrys spp. by LC-MS/MS. Toxins (Basel). 2019; 11(3):133. DOI: 10.3390/toxins11030133 PubMed: 30818881
94. Korpinen EL. Studies on Stachybotrys alternans: IV. Effect of low doses of stachbotrys toxins on pregnancy of mice. Acta Pathol Microbiol Scand B Microbiol Immunol. 1974; 82(4):457-464. PubMed: 4528746
95. Weyel DA, Ellakkani M, Alarie Y, Karol M. An aerosol generator for the resuspension of cotton dust. Toxicol Appl Pharmacol. 1984; 76(3):544-547. DOI: 10.1016/0041-008X(84)90359-4 PubMed: 6506079
96. Frazer DG, Robinson V, DeLong DS, Rose D, Tucker J, Weber KC, Olenchock SA, Jayaraman K. A system for exposing laboratory animals to cotton dust aerosol that is stabilized with feedback control. In: Jacobs RR, Waklyn PJ, editors. Proceedings of the Eleventh Cotton Dust Research Conference, Beltwide Cotton Research Conferences, Dallas, Texas, January 7-8, 1987. Memphis, TN: National Cotton Council; 1987. p. 74-78.
97. Frazer DG, Jones WG, Petsonk EL, Kullman GJ, Barger MW, Afshari A, Jones T, Castranova V. Organic dust exposure from compost handling: Response of an animal model. Am J Ind Med. 1993; 24(4):375-385. DOI: 10.1002/ajim.4700240404 PubMed: 8250058
98. Frazer DG, Robinson VA, Olenchock SA, Hahon N, Barger M, Castranova V. Comparison of the guinea pig's physiological and cellular response to the respirable fraction of untreated, heat treated, microwave irradiated and ethylene oxide treated DB 1/88 bulk cotton dust. In: Jacobs RR, Wakelyn PJ, Domelsmith LN, editors. Proceedings of the Fifteenth Cotton Dust Research Conference, Beltwide Cotton Conferences, San Antonio, Texas, January 11-12, 1991. Memphis, TN: National Cotton Council; 1991. p. 246-251.
99. McKinney W, Chen B, Frazer D. Computer controlled multi-walled carbon nanotube inhalation exposure system. Inhal Toxicol. 2009; 21(12):1053-1061. DOI: 10.1080/08958370802712713 PubMed: 19555230
100. Buskirk AD, Green BJ, Lemons AR, Nayak AP, Goldsmith WT, Kashon ML, Anderson SE, Hettick JM, Templeton SP, Germolec DR, et al. A murine inhalation model to characterize pulmonary exposure to dry Aspergillus fumigatus conidia. PLoS One. 2014; 9(10):e109855. DOI: 10.1371/journal.pone.0109855 PubMed: 25340353
101. Croston TL, Nayak AP, Lemons AR, Goldsmith WT, Gu JK, Germolec DR, Beezhold DH, Green BJ. Influence of Aspergillus fumigatus conidia viability on murine pulmonary microRNA and mRNA expression following subchronic inhalation exposure. Clin Exp Allergy. 2016; 46(10):1315-1327. DOI: 10.1111/cea.12783 PubMed: 27473664
102. Nayak AP, Green BJ, Lemons AR, Marshall NB, Goldsmith WT, Kashon ML, Anderson SE, Germolec DR, Beezhold DH. Subchronic exposures to fungal bioaerosols promotes allergic pulmonary inflammation in naïve mice. Clin Exp Allergy. 2016; 46(6):861-870. DOI: 10.1111/cea.12724 PubMed: 26892490
103. Raabe OG, Al-Bayati MA, Teague SV, Rasolt A. Regional deposition of inhaled monodisperse coarse and fine aerosol particles in small laboratory animals. Ann Occup Hyg. 1988; 32 Suppl 1(Inhaled Particles VI):53-63. DOI: 10.1093/annhyg/32.inhaled_particles_VI.53
104. National Toxicology Program (NTP). Specifications for the conduct of studies to evaluate the toxic and carcinogenic potential of chemical, biological and physical agents in laboratory animals for the National Toxicology Program (NTP). Research Triangle Park, NC: U.S. Department of Health and Human Services, Public Health Service, National Institutes of Health, National Toxicology Program; 2011.
105. Boorman GA, Haseman JK, Waters MD, Hardisty JF, Sills RC. Quality review procedures necessary for rodent pathology databases and toxicogenomic studies: The National Toxicology Program experience. Toxicol Pathol. 2002; 30(1):88-92. DOI: 10.1080/01926230252824752 PubMed: 11890481
106. Maronpot RR, Boorman GA. Interpretation of rodent hepatocellular proliferative alterations and hepatocellular tumors in chemical safety assessment. Toxicol Pathol. 1982; 10(2):71-78. DOI: 10.1177/019262338201000210 PubMed: 28094716
107. Gart JJ, Chu KC, Tarone RE. Statistical issues in interpretation of chronic bioassay tests for carcinogenicity. J Natl Cancer Inst. 1979; 62(4):957-974. PubMed: 285297
108. Tukey JW. Comparing individual means in the analysis of variance. Biometrics. 1949; 5(2):99-114. DOI: 10.2307/3001913 PubMed: 18151955
109. Dunn OJ. Multiple comparisons using rank sums. Technometrics. 1964; 6(3):241-252. DOI: 10.1080/00401706.1964.10490181
110. Dixon WJ, Massey FJ. Introduction to statistical analysis. 2nd ed. New York, NY: McGraw Hill; 1957. p. 145-147, 276-278, 412.
111. U.S. Food and Drug Administration (FDA). 21 CFR Part 58. https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?CFRPart=58
112. National Institute for Occupational Safety and Health (NIOSH). NIOSH Health Effects Laboratory Division (HELD) fact sheet. Cincinnati, OH: U.S. Department of Health and Human Services, Public Health Service, Centers for Disease Control and Prevention, National Institute for Occupational Safety and Health; 2019. DHHS (NIOSH) Publication No. 2019-134. DOI: 10.26616/NIOSHPUB2019134
113. Miller JA, Miller EC. Ultimate chemical carcinogens as reactive mutagenic electrophiles. In: Hiatt HH, Watson JD, Winsten JA, editors. Origins of Human Cancer. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1977. p. 605-627.
114. Straus DS. Somatic mutation, cellular differentiation, and cancer causation. J Natl Cancer Inst. 1981; 67(2):233-241. PubMed: 7021938
115. Crawford BD. Perspectives on the somatic mutation model of carcinogenesis. In: Flamm WG, Lorentzen RJ, editors. Mechanisms and Toxicity of Chemical Carcinogens and Mutagens. Princeton, NJ: Princeton Scientific Publishing; 1985. p. 13-59.
116. Schmid W. The micronucleus test. Mutat Res. 1975; 31(1):9-15. DOI: 10.1016/0165-1161(75)90058-8 PubMed: 48190
117. Heddle JA, Hite M, Kirkhart B, Mavournin K, MacGregor JT, Newell GW, Salamone MF. The induction of micronuclei as a measure of genotoxicity: A report of the U.S. Environmental Protection Agency Gene-Tox Program. Mutat Res. 1983; 123(1):61-118. DOI: 10.1016/0165-1110(83)90047-7 PubMed: 6888413
118. Shelby MD, Erexson GL, Hook GJ, Tice RR. Evaluation of a three-exposure mouse bone marrow micronucleus protocol: Results with 49 chemicals. Environ Mol Mutagen. 1993; 21(2):160-179. DOI: 10.1002/em.2850210210 PubMed: 8444144
119. Shelby MD, Witt KL. Comparison of results from mouse bone marrow chromosome aberration and micronucleus tests. Environ Mol Mutagen. 1995; 25(4):302-313. DOI: 10.1002/em.2850250407 PubMed: 7607185
120. Witt KL, Knapton A, Wehr CM, Hook GJ, Mirsalis J, Shelby MD, MacGregor JT. Micronucleated erythrocyte frequency in peripheral blood of B6C3F(1) mice from short-term, prechronic, and chronic studies of the NTP carcinogenesis bioassay program. Environ Mol Mutagen. 2000; 36(3):163-194. DOI: 10.1002/1098-2280(2000)36:3<163::AID-EM1>3.0.CO;2-P PubMed: 11044899
121. National Toxicology Program (NTP). TOX-107: Pathology tables, survival and growth curves from NTP short-term and genetic toxicology studies. Research Triangle Park, NC: U.S. Department of Health and Human Services, Public Health Service, National Toxicology Program; 2024. DOI: 10.22427/NTP-DATA-TOX-107
122. Li DW, Yang CS. Taxonomic history and current status of Stachybotrys chartarum and related species. Indoor Air. 2005; 15 Suppl 9:5-10. DOI: 10.1111/j.1600-0668.2005.00339.x PubMed: 15910524
123. Roe JD, Haugland RA, Vesper SJ, Wymer LJ. Quantification of Stachybotrys chartarum conidia in indoor dust using real time, fluorescent probe-based detection of PCR products. J Expo Anal Environ Epidemiol. 2001; 11(1):12-20. DOI: 10.1038/sj.jea.7500147 PubMed: 11246797
124. Osimitz TG, Droege W, Finch JM. Toxicologic significance of histologic change in the larynx of the rat following inhalation exposure: A critical review. Toxicol Appl Pharmacol. 2007; 225(3):229-237. DOI: 10.1016/j.taap.2007.08.027 PubMed: 17991503
125. Nagayoshi M, Tada Y, West J, Ochiai E, Watanabe A, Toyotome T, Tanabe N, Takiguchi Y, Shigeta A, Yasuda T, et al. Inhalation of Stachybotrys chartarum evokes pulmonary arterial remodeling in mice, attenuated by Rho-kinase inhibitor. Mycopathologia. 2011; 172(1):5-15. DOI: 10.1007/s11046-011-9400-3 PubMed: 21505873
126. Rosenblum Lichtenstein JH, Molina RM, Donaghey TC, Hsu YHH, Mathews JA, Kasahara DI, Park JA, Bordini A, Godleski JJ, Gillis BS, et al. Repeated mouse lung exposures to Stachybotrys chartarum shift immune response from type 1 to type 2. Am J Respir Cell Mol Biol. 2016; 55(4):521-531. DOI: 10.1165/rcmb.2015-0291OC PubMed: 27148627
127. Leino M, Mäkelä M, Reijula K, Haahtela T, Mussalo-Rauhamaa H, Tuomi T, Hintikka EL, Alenius H. Intranasal exposure to a damp building mould, Stachybotrys chartarum, induces lung inflammation in mice by satratoxin-independent mechanisms. Clin Exp Allergy. 2003; 33(11):1603-1610. DOI: 10.1046/j.1365-2222.2003.01808.x PubMed: 14616875
128. Rao CY, Burge HA, Brain JD. The time course of responses to intratracheally instilled toxic Stachybotrys chartarum spores in rats. Mycopathologia. 2000; 149(1):27-34. DOI: 10.1023/A:1007239017018 PubMed: 11227851
129. Rosenblum Lichtenstein JH, Molina RM, Donaghey TC, Brain JD. Strain differences influence murine pulmonary responses to Stachybotrys chartarum. Am J Respir Cell Mol Biol. 2006; 35(4):415-423. DOI: 10.1165/rcmb.2005-0483OC PubMed: 16690987
130. Innes JR, Ulland BM, Valerio MG, Petrucelli L, Fishbein L, Hart ER, Pallotta AJ, Bates RR, Falk HL, Gart JJ, et al. Bioassay of pesticides and industrial chemicals for tumorigenicity in mice: A preliminary note. J Natl Cancer Inst. 1969; 42(6):1101-1114. PubMed: 5793189
131. Shimodaira K, Okubo Y, Ochiai E, Nakayama H, Katano H, Wakayama M, Shinozaki M, Ishiwatari T, Sasai D, Tochigi N, et al. Gene expression analysis of a murine model with pulmonary vascular remodeling compared to end-stage IPAH lungs. Respir Res. 2012; 13(1):103. DOI: 10.1186/1465-9921-13-103 PubMed: 23157700
132. Hudson B, Flemming J, Sun G, Rand TG. Comparison of immunomodulator mRNA and protein expression in the lungs of Stachybotrys chartarum spore-exposed mice. J Toxicol Environ Health A. 2005; 68(15):1321-1335. DOI: 10.1080/15287390590953572 PubMed: 16020192
133. Rand TG, White K, Logan A, Gregory L. Histological, immunohistochemical and morphometric changes in lung tissue in juvenile mice experimentally exposed to Stachybotrys chartarum spores. Mycopathologia. 2003; 156(2):119-131. DOI: 10.1023/A:1022920205355 PubMed: 12733633
134. Brain JD, Sieber NL, Rosenblum Lichtenstein JH. Killing two birds with one stone: Mold-induced pulmonary immune responses and arterial remodeling. Am J Respir Cell Mol Biol. 2020; 62(5):537-538. DOI: 10.1165/rcmb.2019-0386ED PubMed: 31693387
135. Witt KL, Livanos E, Kissling GE, Torous DK, Caspary W, Tice RR, Recio L. Comparison of flow cytometry-and microscopy-based methods for measuring micronucleated reticulocyte frequencies in rodents treated with nongenotoxic and genotoxic chemicals. Mutat Res. 2008; 649(1-2):101-113. DOI: 10.1016/j.mrgentox.2007.08.004 PubMed: 17869571
Appendices
Appendix A. Culture, Characterization, and Delivery of Stachybotrys chartarum
A.1.
Reconstitution, Culture, and Glycerol Stock Preparation of Stachybotrys chartarum
The Stachybotrys chartarum strain (CBS 414.95, IBT 9460) used in this study was originally isolated from a water-damaged building in Finland. Malt extract agar (MEA) culture plates were aseptically inoculated with 20 µL of the stored glycerol stock in a class II biological safety cabinet (BSC). MEA culture plates were incubated at 26°C for 10–14 days for optimal spore growth. S. chartarum conidia (spores) were harvested by liberating the conidia in 2 mL of sterile water using an inoculating loop. The conidia suspension was mixed with an equal volume of 50% glycerol, and 0.5-mL aliquots of the suspension were prepared in cryogenic storage vials. Glycerol stock preparations were stored at −80°C for the duration of the study.
A.2.
Preparation of the Viable Stachybotrys chartarum Test Article
One aliquoted frozen glycerol conidia stock was thawed, and two MEA culture plates were aseptically inoculated with 20 µL of the stock suspension in a BSC. Plates were incubated at 26°C for 12–14 days. S. chartarum conidia were harvested from each plate by liberating the conidia in 2 mL of sterile water (W3500, Sigma Aldrich, St. Louis, MO) using an inoculating loop. The concentration of the conidia suspension was determined by diluting the suspension 1:100, and conidia counts were determined using a hemocytometer. The conidia suspension was diluted to 2.5 × 105 conidia/mL using sterile water. Mahatma-enriched extra-long grain white rice (30 g) and distilled deionized water (10 mL) were added to twelve 250-mL flasks covered with aluminum foil, autoclaved at 121°C for 30 minutes, and allowed to cool in a BSC. Once cooled, these 12 flasks were aseptically inoculated with 5 mL of the 2.5 × 105 conidia/mL suspension, recapped with foil, and incubated at 26°C for 21–28 days. The flasks were shaken vigorously one time after 1 week of growth to prevent aggregation of the rice grains. At the end of the growth phase, the foil caps were punctured using aseptic technique and the flasks were laid horizontally in a biological safety cabinet to increase the surface area during the drying phase. Cultures were placed in a desiccator for 7–10 days for drying prior to aerosolization. Preparation of the viable test article for the first week of exposures was initiated 1 month before the start of the 3-month exposure period.
A.3.
Preparation of Stachybotrys chartarum Heat-inactivated Particle Control
After 21–28 days of culture, 6 of the 12 fungi-coated rice culture flasks were immersed in a dry bead bath (Lab Armor, Irving, TX) set to 80°C for 2 hours and allowed to cool. As with the viable test articles, the heat-inactivated particle control flasks were then placed in a desiccator for 7–10 days before aerosolization.
A.4.
Determination of the Reduction in Stachybotrys chartarum Conidia Viability in the Heat-inactivated Particle Control
S. chartarum conidia were harvested from heat-inactivated particle control cultures every other week by removing 10–20 random conidia-laden rice grains from the cultures and placing them in a 15-mL tube containing 5 mL sterile water. Conidia suspensions were diluted to 1 × 105 conidia/mL, and 100 µL were plated on MEA culture plates (1 × 104 conidia/plate). After 48–96 hours of growth at 25°C, fungal colony-forming units (CFU) were counted to determine the number of viable S. chartarum conidia/mL. The percentage of viable conidia in the heat-inactivated particle control was then calculated. The cultures were categorized as nonviable if >95% of the conidia were not viable.
A.5.
Assessment of Fungal Contaminants in the Exposure Articles
Genomic DNA was extracted from the heat-inactivated particle control and viable S. chartarum cultures every other week to analyze fungal contaminants using the Roche High Pure PCR Template Preparation Kit (Roche, Indianapolis, IN). Conidia were harvested from 10–20 conidia-laden rice grains in sterile distilled, deionized water. Conidia from 200 µL of the suspension were pelleted by centrifugation and suspended in Roche Tissue Lysis Buffer (200 µL) containing 5 µL of CelLytic B Cell Lysis Reagent (Sigma Aldrich, St. Louis, MO); tubes were incubated at 37°C for 30 minutes. Next, 200 µL Roche Binding Buffer and 40 µL proteinase K solution was added, and tubes were incubated at 70°C for 10 minutes. After the addition of 100 µL of isopropanol, extracted DNA was washed and eluted according to the manufacturer’s recommendations. Internal transcribed spacer (ITS) regions of fungal ribosomal RNA were amplified using the Fun18Sf/ITS4R primer pair, and purified amplicons were cloned into a pDRIVE vector that was then transformed into chemically competent Escherichia coli. E. coli clones positive for the ITS amplicon insert were used to inoculate 96-well plates for the generation of glycerol stocks. The glycerol stocks were sequenced with the T7 and SP6 primers using Sanger sequencing. Forward and reverse sequences were trimmed and assembled and then clustered into operational taxonomic units (groups of sequences with ≥97% similarity). Representative sequences from each operational taxonomic unit were searched against sequences deposited in the National Center for Biotechnology Information database and identified based on sequence identity. Any sequences identified as a fungal species other than S. chartarum or Stachybotrys spp. were considered fungal contaminants. The number of sequences analyzed was dependent on the number of viable conidia in the sample. Very low DNA yield from heat-inactivated particle control samples was a result of heat treatment. The number of contaminant fungal sequences is shown relative to the number of sequences analyzed in each sample in Table A-1.
A.6.
Aerosolization of the Exposure Articles Using the Acoustical Generator System
Prior to aerosolization of the exposure test articles, the viable S. chartarum and heat-inactivated particle control cultures were placed in a desiccator for 7–10 days to remove excess moisture and allow optimal aerosolization. Fungal-laden rice was placed on a rubber membrane housed on top of a speaker within the acoustical generator system (AGS), a modified Pitt-3 generator95 linked to a multianimal nose-only exposure chamber. The Pitt-3 generator was purchased from the University of Pittsburgh and evaluated under conditions recommended by its engineering school.96 The exposure chamber consisted of a vertical cylindrical tower with 24 ports (4 levels of 6 ports) holding animal pods that projected outward radially (Figure 2). The system was fitted with a computer controller, further modified to its current design,97,98 and used by the National Institute for Occupational Safety and Health (NIOSH) to study the respiratory effects caused by the inhalation of a wide variety of occupationally relevant dusts. The AGS was initially used as a new and improved system for dispersing nanomaterials99 and was subsequently modified for fungal spore aerosolization as described by Buskirk et al.100 Two AGS systems were used in this study: one for the viable S. chartarum exposures and one for the heat-inactivated particle control exposures. Air control mice were placed in an identical exposure chamber that was not attached to the AGS but received the same high-efficiency particulate air [filter] (HEPA)-filtered air delivered to the test article chamber. Twenty animal pods were attached to each exposure chamber, 10 for male mice and 10 for female mice. Animals were randomly placed in the pods for each exposure to minimize the effects of potential minor variations in conidia concentrations at each port. The additional ports served as sample ports to collect mass concentration, particle size, and samples on filters for field emission electron microscopy analysis (Hitachi S-4800, Tokyo, Japan).
The fungal-laden rice was allowed to settle within the AGS before the start of exposure. Acoustical energy was then used to aerosolize fungal conidia, which were delivered into the exposure chamber at a flow rate of 6 L/min. Particle size and mass concentration data were needed to estimate the total lung particle deposition during each exposure. Three pods were reserved as sample ports. The first port was used to gravimetrically measure the mass concentration, and the second used a light-scattering device (DataRAM4, ThermoElectron Co., Franklin, MA) to provide real-time estimates of the mass concentration (Con) during exposures. The gravimetric filter data were used to validate and calibrate the DataRAM4 every exposure. The third sample pod was used to collect aerodynamic particle size data (APS, TSI Inc., Soreview, MN). The APS data were collected the week before the start of animal exposures. To collect these data, the system was run in an identical manner as an animal exposure without animals present, and when the real-time mass concentration reading was stable, several 2-minute APS samples were taken. The average DataRam4 readings during the 2-minute APS samples were noted (Con_DuringAPS).
An estimate of the total number of particles deposited in the upper and lower airways was calculated each second (TPDs) by the custom Labview software during each exposure run, using the following equation:
Wherein:
S = deposition scale factor, particle number/(mg/m3)
Con = real-time DataRam4 concentration, mg/m3
The deposition scale factor (S), used in the above equation, was calculated before the beginning of animal exposures with the following equation:
Wherein:
MV = estimated animal breathing minute volume, set to 25 for the current study, mL
i = midpoint of each of the 52 APS aerodynamic size classification channels, ranging from 0.5 to 20 µm
Ncon(i) = the number concentration of particles that fall within the smallest bin size range
DF_Alveolar(i) = mouse lung alveolar deposition fraction for particle size i, ranging from 0 to 1b
DF_Tracheo(i) = mouse lung tracheobronchial deposition fraction for particle size i, ranging from 0 to 1b
Con_DuringAPS = average mass concentration during the APS measurement, mg/m3
The custom Labview software would keep a running total of particles deposited, and when the estimate reached the target daily dose, the particle generator would shut off and deliver HEPA-filtered air to the exposure chamber for the remainder of the 60-minute exposure interval. If the estimated particles deposited did not reach the target within 1 hour, the exposure was stopped, and the estimated count of particles deposited was noted. For the current study, the estimated pulmonary deposition was 1 × 104 conidia, equating to approximately 7 mg/m3 heat-inactivated conidia and 8 mg/m3 viable fungal conidia.
Table A-2 and Table A-3 describe the measured exposure concentrations of each exposure. APS data are shown in Table A-4. The columns represent the three different methods of producing size distribution curves (as seen in Figure 3). The aerodynamic particle sizer totals the particles in each particle size by number, by the surface area of the particles in each size bin, and by the mass of the particles in each size bin. Field emission electron microscopy analysis showed that the aerosol primarily consisted of single conidia within the 3–5 µm range (Figure 3).
bThe mouse-lung-deposition model was based on interpolated data from Raabe et al. 103
Table A-1. Weekly Evaluation of Viable Stachybotrys chartarum and Heat-inactivated Particle Control
| Heat-inactivated Particle Control | Viable S. chartarum | |
|---|---|---|
| Viabilitya | ||
| Viable spore concentration | 1.37 × 103 conidia/mL (±1.62 × 103) |
ND |
| Percentage nonviable | 98.63% (±1.62%) | – |
| Contaminationb | ||
| Week 1 | 0/0c | 0/76 |
| Week 3 | 0/40 | 0/42 |
| Week 5 | 0/39 | 0/93 |
| Week 7 | 0/0c | 0/0c |
| Week 9 | 0/43 | 0/92 |
| Week 11 | 0/0c | 0/85 |
| Week 13 | 0/0c | 0/88 |
Table A-2. Weekly Viable Stachybotrys chartarum Exposure Concentrations
| Exposure Day | Chamber Concentration (DataRAM [mg/m3]) |
Chamber Concentration (Filter [mg/m3]) |
Filter Weight (mg) |
Number of Conidia Deposited in Lung |
|---|---|---|---|---|
| 1 | 8.16 | 8.34 | 0.501 | 1.011 × 104 |
| 3 | 8.13 | 7.32 | 0.440 | 1.009 × 104 |
| 8 | 7.96 | 6.01 | 0.370 | 1.010 × 104 |
| 10 | 8.01 | 7.85 | 0.480 | 1.009 × 104 |
| 15 | 8.10 | 8.54 | 0.512 | 1.008 × 104 |
| 17 | 7.97 | 7.56 | 0.454 | 1.009 × 104 |
| 22 | 8.06 | 8.10 | 0.490 | 1.008 × 104 |
| 24 | 8.03 | 8.20 | 0.500 | 1.008 × 104 |
| 30 | 7.66 | 8.28 | 0.530 | 1.013 × 104 |
| 32 | 8.02 | 10.82 | 0.660 | 1.009 × 104 |
| 36 | 8.12 | 8.49 | 0.510 | 1.008 × 104 |
| 38 | 8.16 | 8.53 | 0.510 | 1.009 × 104 |
| 43 | 7.84 | 8.74 | 0.540 | 1.011 × 104 |
| 45 | 7.94 | 8.02 | 0.497 | 1.010 × 104 |
| 50 | 8.11 | 9.09 | 0.545 | 1.007 × 104 |
| 52 | 8.12 | 10.57 | 0.630 | 1.009 × 104 |
| 58 | 7.62 | 7.58 | 0.485 | 1.008 × 104 |
| 60 | 8.00 | 10.02 | 0.611 | 1.013 × 104 |
| 64 | 7.77 | 9.44 | 0.595 | 1.005 × 104 |
| 66 | 8.10 | 6.87 | 0.412 | 1.009 × 104 |
| 71 | 7.97 | 9.21 | 0.562 | 1.008 × 104 |
| 73 | 8.04 | 8.58 | 0.523 | 1.007 × 104 |
| 78 | 7.50 | 6.90 | 0.449 | 1.009 × 104 |
| 80 | 7.91 | 9.56 | 0.593 | 1.007 × 104 |
| 85 | 8.10 | 9.34 | 0.560 | 1.008 × 104 |
| 87 | 8.01 | 8.97 | 0.547 | 1.009 × 104 |
| Meana | 7.98 ± 0.17 | 8.50 ± 1.12 | 0.520 ± 0.067 | 1.009 × 104 ± 17.51 |
Table A-3. Weekly Heat-inactivated Particle Control Exposure Concentrations
| Exposure Day | Chamber Concentration (DataRAM [mg/m3]) |
Chamber Concentration (Filter [mg/m3]) |
Filter Weight (mg) |
Number of Conidia Deposited in Lung |
|---|---|---|---|---|
| 1 | 7.08 | 6.56 | 0.390 | 1.019 × 104 |
| 3 | 7.10 | 7.54 | 0.450 | 1.023 × 104 |
| 8 | 7.06 | 9.81 | 0.590 | 1.016 × 104 |
| 10 | 7.05 | 8.06 | 0.480 | 1.020 × 104 |
| 15 | 7.13 | 8.66 | 0.515 | 1.028 × 104 |
| 17 | 7.07 | 6.40 | 0.384 | 1.032 × 104 |
| 22 | 7.14 | 9.25 | 0.560 | 1.029 × 104 |
| 24 | 7.15 | 8.70 | 0.520 | 1.027 × 104 |
| 30 | 7.14 | 9.37 | 0.570 | 1.029 × 104 |
| 32 | 7.08 | 8.83 | 0.530 | 1.017 × 104 |
| 36 | 6.80 | 9.59 | 0.590 | 1.011 × 104 |
| 38 | 6.99 | 9.55 | 0.570 | 1.008 × 104 |
| 43 | 7.27 | 9.48 | 0.570 | 1.049 × 104 |
| 45 | 6.96 | 9.77 | 0.596 | 1.019 × 104 |
| 50 | 7.04 | 4.89 | 0.293 | 1.014 × 104 |
| 52 | 7.07 | 6.02 | 0.360 | 1.018 × 104 |
| 58 | 6.94 | 5.78 | 0.353 | 1.021 × 104 |
| 60 | 7.08 | 5.62 | 0.337 | 1.026 × 104 |
| 64 | 7.12 | 7.39 | 0.444 | 1.028 × 104 |
| 66 | 7.10 | 7.10 | NA | 1.026 × 104 |
| 71 | 7.15 | 9.07 | 0.544 | 1.029 × 104 |
| 73 | 7.11 | 8.26 | 0.496 | 1.023 × 104 |
| 78 | 6.79 | 6.04 | 0.374 | 1.017 × 104 |
| 80 | 7.08 | 8.64 | 0.518 | 1.018 × 104 |
| 85 | 7.03 | 8.06 | 0.491 | 1.029 × 104 |
| 87 | 6.99 | 9.41 | 0.574 | 1.028 × 104 |
| Meana | 7.06 ± 0.10 | 7.99 ± 1.50 | 0.484 ± 0.092 | 1.023 × 104 ± 81.84 |
Table A-4. Aerodynamic Particle Size Statistics of Aerosolized Stachybotrys chartarum Based on Aerodynamic Particle Sizer Measurements
| Number (Particle Size) | Surface (Particle Size) | Mass (Particle Size) | |
|---|---|---|---|
| Viable S. chartarum | |||
| Median (µm) | 2.88 ± 0.12 | 4.26 ± 0.03 | 4.43 ± 0.03 |
| Mean (µm) | 2.75 ± 0.05 | 4.14 ± 0.03 | 4.43 ± 0.03 |
| Geometric mean (µm) | 2.22 ± 0.05 | 3.95 ± 0.03 | 4.30 ± 0.02 |
| Mode (µm) | 4.27 ± 0.17 | 4.37 ± 0.00 | 4.37 ± 0.00 |
| Geometric standard deviation | 2.02 ± 0.01 | 1.41 ± 0.01 | 1.29 ± 0.01 |
| Heat-inactivated particle control | |||
| Median (µm) | 3.77 ± 0.17 | 4.10 ± 0.11 | 4.26 ± 0.11 |
| Mean (µm) | 3.77 ± 0.22 | 4.21 ± 0.16 | 4.44 ± 0.20 |
| Geometric mean (µm) | 3.63 ± 0.26 | 4.10 ± 0.16 | 4.32 ± 0.17 |
| Mode (µm) | 3.84 ± 0.11 | 4.02 ± 0.11 | 4.12 ± 0.12 |
| Geometric standard deviation | 1.34 ± 0.07 | 1.25 ± 0.03 | 1.25 ± 0.03 |
Appendix B. Ingredients, Nutrient Composition, and Contaminant Levels in NTP-2000 Mouse Ration
Table B-1. Ingredients of NTP-2000 Mouse Ration
| Ingredients | Percent by Weight |
|---|---|
| Ground hard winter wheat | 23.00 |
| Ground #2 yellow shelled corn | 22.44 |
| Wheat middlings | 15.0 |
| Oat hulls | 8.5 |
| Alfalfa meal (dehydrated, 17% protein) | 7.5 |
| Purified cellulose | 5.5 |
| Soybean meal (49% protein) | 4.0 |
| Fish meal (60% protein) | 4.0 |
| Corn oil (without preservatives) | 3.0 |
| Soy oil (without preservatives) | 3.0 |
| Dried brewer’s yeast | 1.0 |
| Calcium carbonate (USP) | 0.9 |
| Vitamin premixa | 0.5 |
| Mineral premixb | 0.5 |
| Calcium phosphate, dibasic (USP) | 0.4 |
| Sodium chloride | 0.3 |
| Choline chloride (70% choline) | 0.26 |
| Methionine | 0.2 |
Table B-1.. Ingredients of NTP-2000 Mouse Ration
| Ingredients | Percent by Weight |
|---|---|
| Ground hard winter wheat | 23.00 |
| Ground #2 yellow shelled corn | 22.44 |
| Wheat middlings | 15.0 |
| Oat hulls | 8.5 |
| Alfalfa meal (dehydrated, 17% protein) | 7.5 |
| Purified cellulose | 5.5 |
| Soybean meal (49% protein) | 4.0 |
| Fish meal (60% protein) | 4.0 |
| Corn oil (without preservatives) | 3.0 |
| Soy oil (without preservatives) | 3.0 |
| Dried brewer’s yeast | 1.0 |
| Calcium carbonate (USP) | 0.9 |
| Vitamin premixa | 0.5 |
| Mineral premixb | 0.5 |
| Calcium phosphate, dibasic (USP) | 0.4 |
| Sodium chloride | 0.3 |
| Choline chloride (70% choline) | 0.26 |
| Methionine | 0.2 |
Table B-2. Vitamins and Minerals in NTP-2000 Mouse Ration
| Amounta | Source | |
|---|---|---|
| Vitamins | ||
| Vitamin A | 4,000 IU | Stabilized vitamin A palmitate or acetate |
| Vitamin D | 1,000 IU | D-activated animal sterol |
| Vitamin K | 1.0 mg | Menadione sodium bisulfite complex |
| α-Tocopheryl acetate | 100 IU | – |
| Niacin | 23 mg | – |
| Folic acid | 1.1 mg | – |
| α-Pantothenic acid | 10 mg | α-Calcium pantothenate |
| Riboflavin | 3.3 mg | – |
| Thiamine | 4 mg | Thiamine mononitrate |
| B12 | 52 µg | – |
| Pyridoxine | 6.3 mg | Pyridoxine hydrochloride |
| Biotin | 0.2 mg | d-Biotin |
| Minerals | ||
| Magnesium | 514 mg | Magnesium oxide |
| Iron | 35 mg | Iron sulfate |
| Zinc | 12 mg | Zinc oxide |
| Manganese | 10 mg | Manganese oxide |
| Copper | 2.0 mg | Copper sulfate |
| Iodine | 0.2 mg | Calcium iodate |
| Chromium | 0.2 mg | Chromium acetate |
Table B-2.. Vitamins and Minerals in NTP-2000 Mouse Ration
| Amounta | Source | |
|---|---|---|
| Vitamins | ||
| Vitamin A | 4,000 IU | Stabilized vitamin A palmitate or acetate |
| Vitamin D | 1,000 IU | D-activated animal sterol |
| Vitamin K | 1.0 mg | Menadione sodium bisulfite complex |
| α-Tocopheryl acetate | 100 IU | – |
| Niacin | 23 mg | – |
| Folic acid | 1.1 mg | – |
| α-Pantothenic acid | 10 mg | α-Calcium pantothenate |
| Riboflavin | 3.3 mg | – |
| Thiamine | 4 mg | Thiamine mononitrate |
| B12 | 52 µg | – |
| Pyridoxine | 6.3 mg | Pyridoxine hydrochloride |
| Biotin | 0.2 mg | d-Biotin |
| Minerals | ||
| Magnesium | 514 mg | Magnesium oxide |
| Iron | 35 mg | Iron sulfate |
| Zinc | 12 mg | Zinc oxide |
| Manganese | 10 mg | Manganese oxide |
| Copper | 2.0 mg | Copper sulfate |
| Iodine | 0.2 mg | Calcium iodate |
| Chromium | 0.2 mg | Chromium acetate |
Table B-3. Nutrient Composition of NTP-2000 Mouse Ration
| Nutrient | Mean ± Standard Deviation | Range | Number of Samples |
|---|---|---|---|
| Protein (% by weight) | 14.3 | – | 1 |
| Crude fat (% by weight) | 7.9 | – | 1 |
| Crude fiber (% by weight) | 9.49 | – | 1 |
| Ash (% by weight) | 5.05 | – | 1 |
| Amino acids (% of total diet) | |||
| Arginine | 0.808 ± 0.073 | 0.67–0.97 | 31 |
| Cystine | 0.220 ± 0.021 | 0.15–0.25 | 31 |
| Glycine | 0.703 ± 0.037 | 0.62–0.8 | 31 |
| Histidine | 0.341 ± 0.068 | 0.27–0.68 | 31 |
| Isoleucine | 0.548 ± 0.039 | 0.43–0.66 | 31 |
| Leucine | 1.096 ± 0.061 | 0.96–1.24 | 31 |
| Lysine | 0.070 ± 0.101 | 0.31–0.86 | 31 |
| Methionine | 0.409 ± 0.040 | 0.26–0.49 | 31 |
| Phenylalanine | 0.623 ± 0.045 | 0.471–0.72 | 31 |
| Threonine | 0.513 ± 0.040 | 0.43–0.61 | 31 |
| Tryptophan | 0.156 ± 0.026 | 0.11–0.2 | 31 |
| Tyrosine | 0.425 ± 0.064 | 0.28–0.54 | 31 |
| Valine | 0.666 ± 0.038 | 0.55–0.73 | 31 |
| Essential fatty acids (% of total diet) | |||
| Linoleic | 3.936 ± 0.229 | 3.49–4.55 | 31 |
| Linolenic | 0.304 ± 0.030 | 0.21–0.368 | 31 |
| Vitamins | |||
| Vitamin A (IU/kg) | 3,270 | – | 1 |
| α-Tocopherol (ppm) | 2,376 ± 12,602 | 13.6–69,100 | 30 |
| Thiamine (ppm)b | 7.4 | – | 1 |
| Riboflavin (ppm) | 8.32 ± 2.868 | 4.2–17.5 | 31 |
| Niacin (ppm) | 79.78 ± 8.978 | 66.4–98.2 | 31 |
| Pantothenic acid (ppm) | 26.28 ± 10.69 | 17.4–81.0 | 31 |
| Pyridoxine (ppm)b | 9.832 ± 2.080 | 6.44–14.3 | 31 |
| Folic acid (ppm) | 1.61 ± 0.434 | 1.15–3.27 | 31 |
| Biotin (ppm) | 0.319 ± 0.113 | 0.0–0.704 | 31 |
| B12 (ppb) | 48.82 ± 33.79 | 18.3–174.0 | 31 |
| Choline (as chloride) (ppm) | 2,553 ± 632 | 1,160–3,790 | 31 |
| Minerals | |||
| Calcium (%) | 0.943 | – | 1 |
| Phosphorus (%) | 0.573 | – | 1 |
| Potassium (%) | 0.663 ± 0.035 | 0.563–0.733 | 31 |
| Chloride (%) | 0.389 ± 0.044 | 0.3–0.517 | 31 |
| Sodium (%) | 0.194 ± 0.027 | 0.153–0.283 | 31 |
| Magnesium (%) | 0.216 ± 0.052 | 0.185–0.49 | 31 |
| Iron (ppm) | 190.0 ± 35.69 | 135–311 | 31 |
| Manganese (ppm) | 49.87 ± 9.15 | 21.0–73.1 | 31 |
| Zinc (ppm) | 56.53 ± 24.87 | 42.5–184 | 31 |
| Copper (ppm) | 7.64 ± 2.42 | 3.21–16.3 | 31 |
| Iodine (ppm) | 0.50 ± 0.232 | 0–0.972 | 31 |
| Chromium (ppm) | 1.164 ± 1.16 | 0.33–3.97 | 30 |
| Cobalt (ppm) | 0.217 ± 0.148 | 0.086–0.864 | 29 |
Table B-3.. Nutrient Composition of NTP-2000 Mouse Ration
| Nutrient | Mean ± Standard Deviation | Range | Number of Samples |
|---|---|---|---|
| Protein (% by weight) | 14.3 | – | 1 |
| Crude fat (% by weight) | 7.9 | – | 1 |
| Crude fiber (% by weight) | 9.49 | – | 1 |
| Ash (% by weight) | 5.05 | – | 1 |
| Amino acids (% of total diet) | |||
| Arginine | 0.808 ± 0.073 | 0.67–0.97 | 31 |
| Cystine | 0.220 ± 0.021 | 0.15–0.25 | 31 |
| Glycine | 0.703 ± 0.037 | 0.62–0.8 | 31 |
| Histidine | 0.341 ± 0.068 | 0.27–0.68 | 31 |
| Isoleucine | 0.548 ± 0.039 | 0.43–0.66 | 31 |
| Leucine | 1.096 ± 0.061 | 0.96–1.24 | 31 |
| Lysine | 0.070 ± 0.101 | 0.31–0.86 | 31 |
| Methionine | 0.409 ± 0.040 | 0.26–0.49 | 31 |
| Phenylalanine | 0.623 ± 0.045 | 0.471–0.72 | 31 |
| Threonine | 0.513 ± 0.040 | 0.43–0.61 | 31 |
| Tryptophan | 0.156 ± 0.026 | 0.11–0.2 | 31 |
| Tyrosine | 0.425 ± 0.064 | 0.28–0.54 | 31 |
| Valine | 0.666 ± 0.038 | 0.55–0.73 | 31 |
| Essential fatty acids (% of total diet) | |||
| Linoleic | 3.936 ± 0.229 | 3.49–4.55 | 31 |
| Linolenic | 0.304 ± 0.030 | 0.21–0.368 | 31 |
| Vitamins | |||
| Vitamin A (IU/kg) | 3,270 | – | 1 |
| α-Tocopherol (ppm) | 2,376 ± 12,602 | 13.6–69,100 | 30 |
| Thiamine (ppm)b | 7.4 | – | 1 |
| Riboflavin (ppm) | 8.32 ± 2.868 | 4.2–17.5 | 31 |
| Niacin (ppm) | 79.78 ± 8.978 | 66.4–98.2 | 31 |
| Pantothenic acid (ppm) | 26.28 ± 10.69 | 17.4–81.0 | 31 |
| Pyridoxine (ppm)b | 9.832 ± 2.080 | 6.44–14.3 | 31 |
| Folic acid (ppm) | 1.61 ± 0.434 | 1.15–3.27 | 31 |
| Biotin (ppm) | 0.319 ± 0.113 | 0.0–0.704 | 31 |
| B12 (ppb) | 48.82 ± 33.79 | 18.3–174.0 | 31 |
| Choline (as chloride) (ppm) | 2,553 ± 632 | 1,160–3,790 | 31 |
| Minerals | |||
| Calcium (%) | 0.943 | – | 1 |
| Phosphorus (%) | 0.573 | – | 1 |
| Potassium (%) | 0.663 ± 0.035 | 0.563–0.733 | 31 |
| Chloride (%) | 0.389 ± 0.044 | 0.3–0.517 | 31 |
| Sodium (%) | 0.194 ± 0.027 | 0.153–0.283 | 31 |
| Magnesium (%) | 0.216 ± 0.052 | 0.185–0.49 | 31 |
| Iron (ppm) | 190.0 ± 35.69 | 135–311 | 31 |
| Manganese (ppm) | 49.87 ± 9.15 | 21.0–73.1 | 31 |
| Zinc (ppm) | 56.53 ± 24.87 | 42.5–184 | 31 |
| Copper (ppm) | 7.64 ± 2.42 | 3.21–16.3 | 31 |
| Iodine (ppm) | 0.50 ± 0.232 | 0–0.972 | 31 |
| Chromium (ppm) | 1.164 ± 1.16 | 0.33–3.97 | 30 |
| Cobalt (ppm) | 0.217 ± 0.148 | 0.086–0.864 | 29 |
Table B-4. Contaminant Levels in NTP-2000 Mouse Ration
| Mean | Number of Samples | |
|---|---|---|
| Contaminants | ||
| Arsenic (ppm) | 0.252 | 1 |
| Cadmium (ppm) | 0.051 | 1 |
| Lead (ppm) | 0.066 | 1 |
| Mercury (ppm) | 0.01 | 1 |
| Selenium (ppm) | 0.145 | 1 |
| Aflatoxins (ppb)a | <5.0 | 1 |
| Nitrate nitrogen (ppm)b | 11.5 | 1 |
| Nitrite nitrogen (ppm)a,b | <0.12 | 1 |
| BHA (ppm)a,c | <1.0 | 1 |
| BHT (ppm)a,c | <1.0 | 1 |
| Aerobic plate count (CFU/g)a | <10.0 | 1 |
| Coliform (MPN/g)a | <3 | 1 |
| Escherichia coli (MPN/g)a | <10 | 1 |
| Salmonella sp. (MPN/g) | Negative | 1 |
| Total nitrosamines (ppb)d | 9.1 | 1 |
| N-Nitrosodimethylamine (ppb)d | 5.0 | 1 |
| N-Nitrosopyrrolidine (ppb)d | 4.1 | 1 |
| Pesticides (ppm) | ||
| α-BHCa | <0.01 | 1 |
| β-BHCa | <0.02 | 1 |
| γ-BHCa | <0.01 | 1 |
| δ-BHCa | <0.01 | 1 |
| Heptachlora | <0.01 | 1 |
| Aldrina | <0.01 | 1 |
| Heptachlor epoxidea | <0.01 | 1 |
| DDEa | <0.01 | 1 |
| DDDa | <0.01 | 1 |
| DDTa | <0.01 | 1 |
| HCBa | <0.01 | 1 |
| Mirexa | <0.01 | 1 |
| Methoxychlora | <0.05 | 1 |
| Dieldrina | <0.01 | 1 |
| Endrina | <0.01 | 1 |
| Telodrina | <0.01 | 1 |
| Chlordanea | <0.05 | 1 |
| Toxaphenea | <0.01 | 1 |
| Estimated PCBsa | <0.20 | 1 |
| Ronnela | <0.01 | 1 |
| Ethiona | <0.02 | 1 |
| Trithiona | <0.05 | 1 |
| Diazinona | <0.10 | 1 |
| Methyl chlorpyrifos | 0.19 | 1 |
| Ethyl chlorpyrifos | <0.025 | 1 |
| Methyl pirimiphos | <0.02 | 1 |
| Methyl parathiona | <0.025 | 1 |
| Ethyl parathiona | <0.02 | 1 |
| Malathiona | <0.02 | 1 |
| Endosulfan Ia | <0.02 | 1 |
| Endosulfan II | 0.12 | 1 |
| Endosulfane sulfatea | <0.01 | 1 |
Table B-4.. Contaminant Levels in NTP-2000 Mouse Ration
| Mean | Number of Samples | |
|---|---|---|
| Contaminants | ||
| Arsenic (ppm) | 0.252 | 1 |
| Cadmium (ppm) | 0.051 | 1 |
| Lead (ppm) | 0.066 | 1 |
| Mercury (ppm) | 0.01 | 1 |
| Selenium (ppm) | 0.145 | 1 |
| Aflatoxins (ppb)a | <5.0 | 1 |
| Nitrate nitrogen (ppm)b | 11.5 | 1 |
| Nitrite nitrogen (ppm)a,b | <0.12 | 1 |
| BHA (ppm)a,c | <1.0 | 1 |
| BHT (ppm)a,c | <1.0 | 1 |
| Aerobic plate count (CFU/g)a | <10.0 | 1 |
| Coliform (MPN/g)a | <3 | 1 |
| Escherichia coli (MPN/g)a | <10 | 1 |
| Salmonella sp. (MPN/g) | Negative | 1 |
| Total nitrosamines (ppb)d | 9.1 | 1 |
| N-Nitrosodimethylamine (ppb)d | 5.0 | 1 |
| N-Nitrosopyrrolidine (ppb)d | 4.1 | 1 |
| Pesticides (ppm) | ||
| α-BHCa | <0.01 | 1 |
| β-BHCa | <0.02 | 1 |
| γ-BHCa | <0.01 | 1 |
| δ-BHCa | <0.01 | 1 |
| Heptachlora | <0.01 | 1 |
| Aldrina | <0.01 | 1 |
| Heptachlor epoxidea | <0.01 | 1 |
| DDEa | <0.01 | 1 |
| DDDa | <0.01 | 1 |
| DDTa | <0.01 | 1 |
| HCBa | <0.01 | 1 |
| Mirexa | <0.01 | 1 |
| Methoxychlora | <0.05 | 1 |
| Dieldrina | <0.01 | 1 |
| Endrina | <0.01 | 1 |
| Telodrina | <0.01 | 1 |
| Chlordanea | <0.05 | 1 |
| Toxaphenea | <0.01 | 1 |
| Estimated PCBsa | <0.20 | 1 |
| Ronnela | <0.01 | 1 |
| Ethiona | <0.02 | 1 |
| Trithiona | <0.05 | 1 |
| Diazinona | <0.10 | 1 |
| Methyl chlorpyrifos | 0.19 | 1 |
| Ethyl chlorpyrifos | <0.025 | 1 |
| Methyl pirimiphos | <0.02 | 1 |
| Methyl parathiona | <0.025 | 1 |
| Ethyl parathiona | <0.02 | 1 |
| Malathiona | <0.02 | 1 |
| Endosulfan Ia | <0.02 | 1 |
| Endosulfan II | 0.12 | 1 |
| Endosulfane sulfatea | <0.01 | 1 |
Appendix C. National Institute for Occupational Safety and Health Sentinel Animal Program
C.1.
Methods
The sentinel animal program at the National Institute for Occupational Safety and Health (NIOSH) monitors for common rodent pathogens in each study room to ensure study mice are healthy and pathogen free. Pathogens are monitored through the collection of blood and feces from sentinel mice housed in the study rooms.
For this toxicity study, mice weighing >10% or <10% of the mean weight of all male or female study mice were selected for the sentinel group (n = 20; 10/sex). Females were group housed with five per cage. Male mice were single housed. For each week the mice were on study, sentinel animals were exposed to dirty bedding from study animals. The 4-week sentinels were evaluated for viral antibodies, Helicobacter, and pinworms 24 hours following the second exposure in the fourth exposure week. The 3-month sentinels were evaluated only at study termination. All sentinels were euthanized at study termination and evaluated for abnormalities by gross necropsy.
Whole blood was collected via tail vein bleed for 4-week sentinels or via cardiac puncture postmortem for 3-month sentinels. Three to four drops of whole blood were placed onto an Opti-Spot card (IDEXX Laboratories, Inc., Westbrook, ME) for serology tests. These cards were submitted to IDEXX BioResearch for analysis using the Opti-HM serology profile. The profile included Mycoplasma pulmonis, ectromelia virus, epizootic diarrhea of infant mice virus, lymphocytic choriomeningitis virus, mouse hepatitis virus, minute virus of mice, mouse parvovirus, mouse norovirus, pneumonia virus of mice, reovirus type 3, Sendai virus, and Theiler’s murine encephalomyelitis virus.
One fecal pellet from each sentinel mouse was collected with sterile forceps, and all pellets for each sample group were placed in a single sterile sample tube for analysis (maximum of 10 pellets/tube). Samples were submitted to IDEXX BioResearch for polymerase chain reaction (PCR) analysis using the Helicobacter profile. The profile included Helicobacter bili, H. ganmani, H. hepaticus, H. mastomyrinus, H. rodentium, H. typhlonius, and Helicobacter spp. Fur pelt swabs were also collected from each 4-week sentinel mouse and submitted to IDEXX BioResearch for PCR evaluation of the pinworms Aspiculuris tetraptera and Syphacia obvelata. Swabs were pooled for the analysis.
C.2.
Results
All test results were negative (Table C-1).
Table C-1. Methods and Results for Sentinel Animal Testing in Male and Female Mice
| Three-month Study | ||
|---|---|---|
| Collection Time Points | 4 Weeks | Study Termination |
| Number examined (males/females) | 5/5 | 5/5 |
| Method/test | ||
| Multiplex fluorescent immunoassay (MFI)a | ||
| Ectromelia virus | – | – |
| Epizootic diarrhea of infant mice (EDIM) |
– | – |
| Lymphocytic choriomeningitis virus | – | – |
| Mycoplasma pulmonis | – | – |
| Mouse hepatitis virus (MHV) | – | – |
| Mouse norovirus (MNV) | – | – |
| Mouse parvovirus (MPV) | – | – |
| Minute virus of mice (MVM) | – | – |
| Pneumonia virus of mice (PVM) | – | – |
| Reovirus type 3 (REO3) | – | – |
| Sendai | – | – |
| Theiler’s murine encephalomyelitis virus (TMEV) GDVII |
– | – |
| PCR evaluation | ||
| Helicobacterb | – | – |
| Pinwormsc | – | NT |
Appendix D. Genetic Toxicology
D.1.
Evaluation Protocol
National Toxicology Program (NTP) reports consider biological as well as statistical factors to determine an overall assay result. For an individual assay, the statistical procedures for data analysis are described in the following protocols. There have been instances, however, in which multiple samples of a chemical were tested in the same assay, and different results were obtained among these samples and/or among laboratories. In such cases, all the data are critically evaluated with attention given to possible protocol variations in determining the weight of evidence for an overall conclusion of chemical activity in an assay. For in vitro assays conducted with and without exogenous metabolic activation, results obtained in the absence of activation are analyzed separately from results obtained in the presence of activation. The summary table in the abstract of this Toxicity Report presents the scientific judgment of the Division of Translational Toxicology (DTT) and the National Institute for Occupational Safety and Health (NIOSH) regarding the overall evidence for activity of the chemical in an assay.
D.2.
Micronucleus Assay
D.2.1.
Peripheral Blood Micronucleus Test Protocol
Peripheral blood samples were analyzed by Integrated Laboratory Systems, LLC (ILS; Research Triangle Park, NC) for determination of erythrocyte micronucleus frequencies. Briefly, ethylenediaminetetraacetic acid (EDTA)-stabilized peripheral blood samples were shipped on ice packs immediately following terminal necropsy from NIOSH to ILS. Before ILS received the samples, microcentrifuge tubes containing anticoagulant (heparin) were prepared and stored at 4°C, and 15 mL conical tubes containing fixative (methanol) were prepared and stored at −80°C ± 5°C.
Upon arrival, blood samples were fixed in ultracold methanol using a MicroFlowPLUS Kit (Litron Laboratories, Rochester, NY) according to the manufacturer’s instructions. Fixed samples were stored in a −80°C freezer until analysis. Thawed blood samples were analyzed for frequency of micronucleated immature erythrocytes (polychromatic erythrocytes or PCEs, reticulocytes) and mature erythrocytes (normochromatic erythrocytes or NCEs) using a flow cytometer135; both the mature and immature erythrocyte populations can be analyzed separately by employing special cell surface markers to differentiate the two cell types. In mice, both the mature and immature erythrocyte populations can be evaluated for micronucleus frequency because the mouse spleen does not sequester and eliminate damaged erythrocytes. Damaged erythrocytes achieve steady state in the peripheral blood of mice following 4 weeks of continuous exposure. Approximately 20,000 reticulocytes and 1 × 106 erythrocytes were analyzed per animal for the frequency of micronucleated cells, and the percentage of immature erythrocytes (% PCE) was calculated as a measure of bone marrow toxicity resulting from test article exposure.
For most NTP studies, the statistical methods for micronucleus assays employ a trend test followed by Dunn pairwise tests. Due to the design of this study (with one exposed group and two control groups), no trend test is needed, and the Dunn test was used for the three-way pairwise comparison of the control and exposed groups. The Dunn test has a built-in correction for multiple comparisons so the overall false positive rate of the experiment can be maintained. Pairwise comparisons with the control groups are considered statistically significant at p ≤ 0.05.
Historical control data are used to evaluate the biological importance of any observed response. Both statistical significance and biological importance are considered when arriving at a call. The results of statistical analyses, reproducibility of any effects observed, and the magnitudes of those effects are all considered when determining the final call.
D.2.2.
Results
No significant increases in the frequencies of micronucleated reticulocytes and mature erythrocytes were observed in the peripheral blood of male or female B6C3F1/N mice exposed to either viable S. chartarum or heat-inactivated particle control conidia relative to the air control mice (Table D-1). Similarly, no significant increase in frequency was observed in mice exposed to viable S. chartarum conidia compared to mice exposed to the heat-inactivated particle control. In addition, no significant alteration in the percentage of reticulocytes was observed in these mice, suggesting no effects on hematopoiesis. These negative results in the micronucleus test indicate an absence of exposure-induced chromosomal damage in progenitor erythrocytes in the bone marrow of mice, along with an absence of generalized bone marrow toxicity.
Table D-1. Frequency of Micronuclei in Peripheral Blood Erythrocytes of Male and Female Mice in the Three-month Inhalation Study of Stachybotrys chartarum
| Number of Mice with Erythrocytes Scored | MN-RET/1,000a | P Valueb | MN-ME/1,000a | P Valueb | % RETa | P Valueb | |
|---|---|---|---|---|---|---|---|
| Male | |||||||
| Air control | 5 | 3.120 ± 0.12 | 1.636 ± 0.04 | 1.844 ± 0.06 | |||
| Heat-inactivated particle control | 5 | 3.200 ± 0.37 | 1.000 | 1.672 ± 0.05 | 1.000 | 1.769 ± 0.09 | 0.688 |
| Viable S. chartarumc | 5 | 3.040 ± 0.12 | 1.000/1.000 | 1.618 ± 0.08 | 1.000/0.688 | 1.764 ± 0.08 | 1.000/1.000 |
| Female | |||||||
| Air control | 5 | 1.820 ± 0.15 | 1.027 ± 0.03 | 1.900 ± 0.10 | |||
| Heat-inactivated particle control | 5 | 2.120 ± 0.18 | 0.384 | 1.084 ± 0.04 | 0.867 | 2.031 ± 0.07 | 1.000 |
| Viable S. chartarumc | 5 | 1.960 ± 0.11 | 1.000/1.000 | 1.004 ± 0.05 | 1.000/0.609 | 1.954 ± 0.21 | 1.000/1.000 |
Appendix E. Supplemental Data
Tables with supplemental data can be found here: https://doi.org/10.22427/NTP-DATA-TOX-107.121
About This Report
Foreword
The National Toxicology Program (NTP), established in 1978, is an interagency program within the Public Health Service of the U.S. Department of Health and Human Services. Its activities are executed through a partnership of the National Institute for Occupational Safety and Health (NIOSH, part of the Centers for Disease Control and Prevention), the Food and Drug Administration (FDA, primarily at the National Center for Toxicological Research), and the National Institute of Environmental Health Sciences (NIEHS, part of the National Institutes of Health), where the program is administratively located. NTP offers a unique venue for the testing, research, and analysis of agents of concern to identify toxic and biological effects, provide information that strengthens the science base, and inform decisions by health regulatory and research agencies to safeguard public health. NTP also works to develop and apply new and improved methods and approaches that advance toxicology and better assess health effects from environmental exposures.
The Toxicity Report series began in 1991. The studies described in the NTP Toxicity Report series are designed and conducted to characterize and evaluate the toxicological potential of selected substances in laboratory animals (usually two species, rats and mice). Substances (e.g., chemicals, physical agents, and mixtures) selected for NTP toxicity studies are chosen primarily on the basis of human exposure, level of commercial production, and chemical structure. The interpretive conclusions presented in the Toxicity Reports are derived solely from the results of these NTP studies and should not be misconstrued to represent an official policy of the individual agencies that participate in the NTP partnership (NIEHS, NIOSH, or FDA). Extrapolation of these results to other species, including characterization of hazards and risks to humans, requires analyses beyond the intent of these reports. Selection for study per se is not an indicator of a substance’s toxic potential.
NTP conducts its studies in compliance with its laboratory health and safety guidelines and Food and Drug Administration Good Laboratory Practice Regulations and meets or exceeds all applicable federal, state, and local health and safety regulations. Animal care and use are in accordance with the Public Health Service Policy on Humane Care and Use of Laboratory Animals. Studies are subjected to retrospective quality assurance audits before they are presented for public review. Draft reports undergo external peer review before they are finalized and published.
NTP Toxicity Reports are available free of charge on the NTP website and cataloged in PubMed, a free resource developed and maintained by the National Library of Medicine (part of the National Institutes of Health). Data for these studies are included in NTP’s Chemical Effects in Biological Systems database.
For questions about the reports and studies, please email NTP or call 984-287-3211.
Collaborators
Division of Translational Toxicology, National Institute of Environmental Health Sciences, Research Triangle Park, North Carolina, USA
Designed study, evaluated and interpreted results, and reported findings
D.R. Germolec, Ph.D., Lead Scientist
C.R. Blystone, Ph.D.
M.F. Cesta, D.V.M., Ph.D.
B.J. Collins, M.S.P.H.
M.C. Cora, D.V.M.
H.C. Cunny, Ph.D.
M.J. Hooth, Ph.D.
A.P. King-Herbert, D.V.M.
D.E. Malarkey, D.V.M., Ph.D. (Retired)
G.K. Roberts, Ph.D.
K.A. Shipkowski, Ph.D.
K.R. Shockley, Ph.D.
S.L. Smith-Roe, Ph.D.
M.D. Stout, Ph.D.
G. Travlos, Ph.D.
N.J. Walker, Ph.D.
Provided oversight for data management
J.M. Fostel, Ph.D.
Integrated Laboratory Systems, LLC, an Inotiv Company, Research Triangle Park, North Carolina, USA
Evaluated and interpreted results and reported findings
C.J. Willson, D.V.M., Ph.D., Lead Pathologist
Coordinated pathology data review and coordinated Pathology Working Group (3-month study) (October 25, 2022)
R.R. Moore, D.V.M.
National Institute for Occupational Safety and Health, Morgantown, West Virginia, USA
Developed test article and exposure system, conducted study, evaluated and interpreted results, and reported findings
D.H. Beezhold, Ph.D., Co-Principal Investigator (Retired)
T.L. Croston, Ph.D., Co-Principal Investigator
B.J. Green, Ph.D., Co-Principal Investigator
L. Battelli, M.S.
W.T. Goldsmith, B.S.
B.F. Law, M.S.
A.R. Lemons, M.S.
W.G. McKinney, M.S.
A.P. Nayak, Ph.D.
M.S. Orandle, D.V.M., Ph.D.
Battelle, Columbus, Ohio, USA
Evaluated pathology findings
K.A.B. Knostman, D.V.M., Ph.D.
K.M. Patton, D.V.M., Ph.D.
ICF, Reston, Virginia, USA
Contributed to technical writing and data integration and ensured report quality
S.A. Gillera, Ph.D.
L.M. Prince, Ph.D.
Contributors
Division of Translational Toxicology, National Institute of Environmental Health Sciences, Research Triangle Park, North Carolina, USA
Provided oversight of external peer review
M.L. Brownlow, Ph.D.
M.S. Wolfe, Ph.D.
Pathology Working Group, National Institute of Environmental Health Sciences, Research Triangle Park, North Carolina, USA
Participated in Pathology Working Group on 3-month mice (October 25, 2022)
M.F. Cesta, D.V.M., Ph.D., National Institute of Environmental Health Sciences
V.S. Chen, D.V.M., Ph.D., Charles River Laboratories, Inc.
K.Y. Cimon, D.V.M., M.S., Experimental Pathology Laboratories, Inc.
T.A. Crabbs, D.V.M., Experimental Pathology Laboratories, Inc.
M.R. Elwell, D.V.M., Ph.D., Consultant
R.R. Moore, D.V.M., Integrated Laboratory Systems, LLC, an Inotiv Company
C.J. Willson, D.V.M., Ph.D., Integrated Laboratory Systems, LLC, an Inotiv Company
Integrated Laboratory Systems, LLC, an Inotiv Company, Research Triangle Park, North Carolina, USA
Supervised pathology review
G.D. Hill, D.V.M., Ph.D.
Conducted micronucleus assay
L. Recio, Ph.D., Principal Investigator
C.A. Hobbs, Ph.D.
CSS Corporation, Research Triangle Park, North Carolina, USA
Prepared quality assessment audits
S. Brecher, Ph.D., Principal Investigator
H. Dimig, B.S.
S. Iyer, B.S.
Social & Scientific Systems, a DLH Company, Research Triangle Park, North Carolina, USA
Provided statistical analyses
S.J. McBride, Ph.D., Principal Investigator
L.J. Betz, M.S.
S.F. Harris, M.S.
A. Jeffers, B.S.
G. Xie, Ph.D.
ASRC Federal, Research Triangle Park, North Carolina, USA
Prepared data for report
J. Berke, B.S.
P. Brown, B.S.
K. Gilbert, B.S.
C. Myers, M.S.
L. Yang, Ph.D.
ICF, Reston, Virginia, USA
Provided contract oversight
D. Burch, M.E.M., Principal Investigator
J.A. Wignall, M.S.P.H.
Provided program oversight
C.E. Haver, M.P.H.
Prepared and edited report
K.S. Duke, Ph.D.
J.I. Powers, M.A.P.
K.T. O’Donovan, B.A.
S.J. Snow, Ph.D.
N. Ukpabi, M.S.
Supported external peer review
L.M. Green, M.P.H
T. Dawson
Peer Review
The National Toxicology Program (NTP) conducted a peer review of the draft NTP Technical Report on the Toxicity Study of Stachybotrys chartarum (CASRN 67892-26-6) Administered by Inhalation to B6C3F1/N Mice by letter in January 2024 by the experts listed below. Reviewer selection and document review followed established NTP practices. The reviewers were charged to:
-
Peer review the draft NTP Technical Report on the Toxicity Study of Stachybotrys chartarum (CASRN 67892-26-6) Administered by Inhalation to B6C3F1/N Mice.
-
Comment on NTP’s interpretations of the data.
NTP carefully considered reviewer comments in finalizing this report.
Peer Reviewers
Kara Corps, D.V.M., Ph.D.
Assistant Professor, College of Veterinary Medicine
The Ohio State University
Columbus, Ohio, USA
James Pestka, Ph.D.
University Distinguished Professor, Department of Food Science and Human Nutrition, Department of Microbiology and Molecular Genetics, Institute for Integrative Toxicology
Michigan State University
East Lansing, Michigan, USA
Publication Details
Publisher: National Toxicology Program
Publishing Location: Research Triangle Park, NC
ISSN: 2378-8992
DOI: https://doi.org/10.22427/NTP-TOX-107
Report Series: NTP Toxicity Report Series
Report Series Number: 107
Official citation: National Toxicology Program (NTP). 2024. NTP technical report on the toxicity study of Stachybotrys chartarum (CASRN 67892-26-6) administered by inhalation to B6C3F1/N mice. Research Triangle Park, NC: National Toxicology Program. Toxicity Report 107.
Acknowledgments
The NIOSH co-authorship team would like to personally thank the NIOSH Inhalation Facility staff for weighing the animals and conducting the inhalation exposures of test article, heat-inactivated biological particle control, and air control groups. The NIOSH co-authorship team would also like to thank the NIOSH animal facility staff for ordering animals and diet, daily clinical observations, and overseeing the sentinel mice housing and testing program. This study was supported by internal NIOSH funds and in part by an interagency agreement between NIOSH and the National Institute of Environmental Health Sciences as a collaborative National Toxicology Program research activity (AES12007001-1-0-6-NIOSH reference 12-NS12-01).
This work was supported by the Intramural Research Program (ES103374, ES103376, ES103377, and ES103380) at the National Institute of Environmental Health Sciences, National Institutes of Health and performed for the National Toxicology Program, Public Health Service, U.S. Department of Health and Human Services under contracts HHSN273201800006C, GS00Q14OADU417 (Order No. HHSN273201600015U), HHSN273201600011C, HHSN273201600020C, HHSN273201500006C, HHSN273201500013C, HHSN273201300009C, HHSN273201300004C, HHSN316201200054W, and N01-ES-55536.