

Nervous System
Brain
Narrative
Recent changes in neuropathology evaluation in NTP studies include routine examination of seven sections of the brain instead of the traditional three-section approach. The modified seven sections include the traditional three sections and four additional sections that include neuroanatomic areas and subsites such as the olfactory bulb, superior colliculus, medial geniculate body, inferior colliculus, substantia nigra, and area postrema, to name a few. In fact, approximately 50 neuroanatomic sites will be available for routine evaluation with the NTP seven-section approach. The rationale and basis for the seven-section approach are described in Rao et al. (2011).
Figure 1 outlines the landmarks for the seven sections in the rat. The traditional three-section approach included levels 2, 3, and 6. The modified approach now includes the addition of four sections (levels 1, 4, 5, and 7), for a total of seven sections.
The neuroanatomy and functional significance of the ~50 neuroanatomic sites in the NTP-7 are detailed in Rao et al. (in press). The Society of Toxicologic Pathology best practices approach for neuropathologic evaluations also includes a seven-section approach as detailed by Bolon et al. (2013). Although not identical, the trimming planes are comparable to the planes depicted in Rao et al, 2011 and Bolon et al, 2013. It is important to note that modest variations in trimming planes are expected variations in brain sampling among animals within a single study, across multiple studies, and among institutions.
This introduction to the compilation of non-neoplastic lesions of the central nervous system lists commonly encountered neuroanatomic structures that are normal and should not be diagnosed as lesions. In addition, commonly encountered artifacts such as basophilic/dark neurons and white matter vacuolation resulting from poor fixation and handling procedures are described.
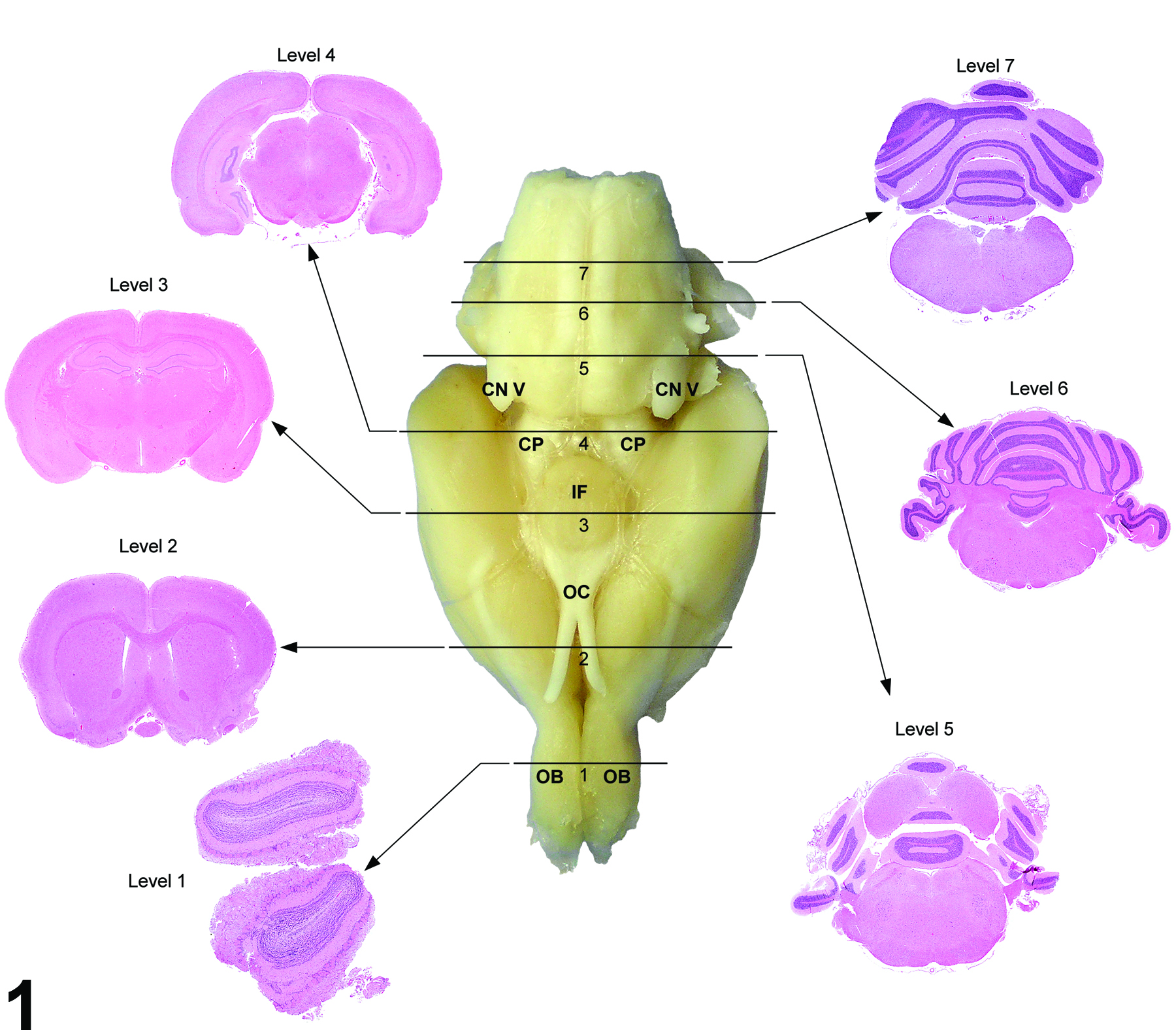
Figure 1. Seven transverse (coronal) hematoxylin and eosin-stained sections corresponding to levels based on anatomic target landmarks. OB = olfactory bulb, OC = optic chiasm, IF = infundibulum (and/or median eminence), CN V = cranial nerve V (trigeminal), CP = cerebral peduncle.
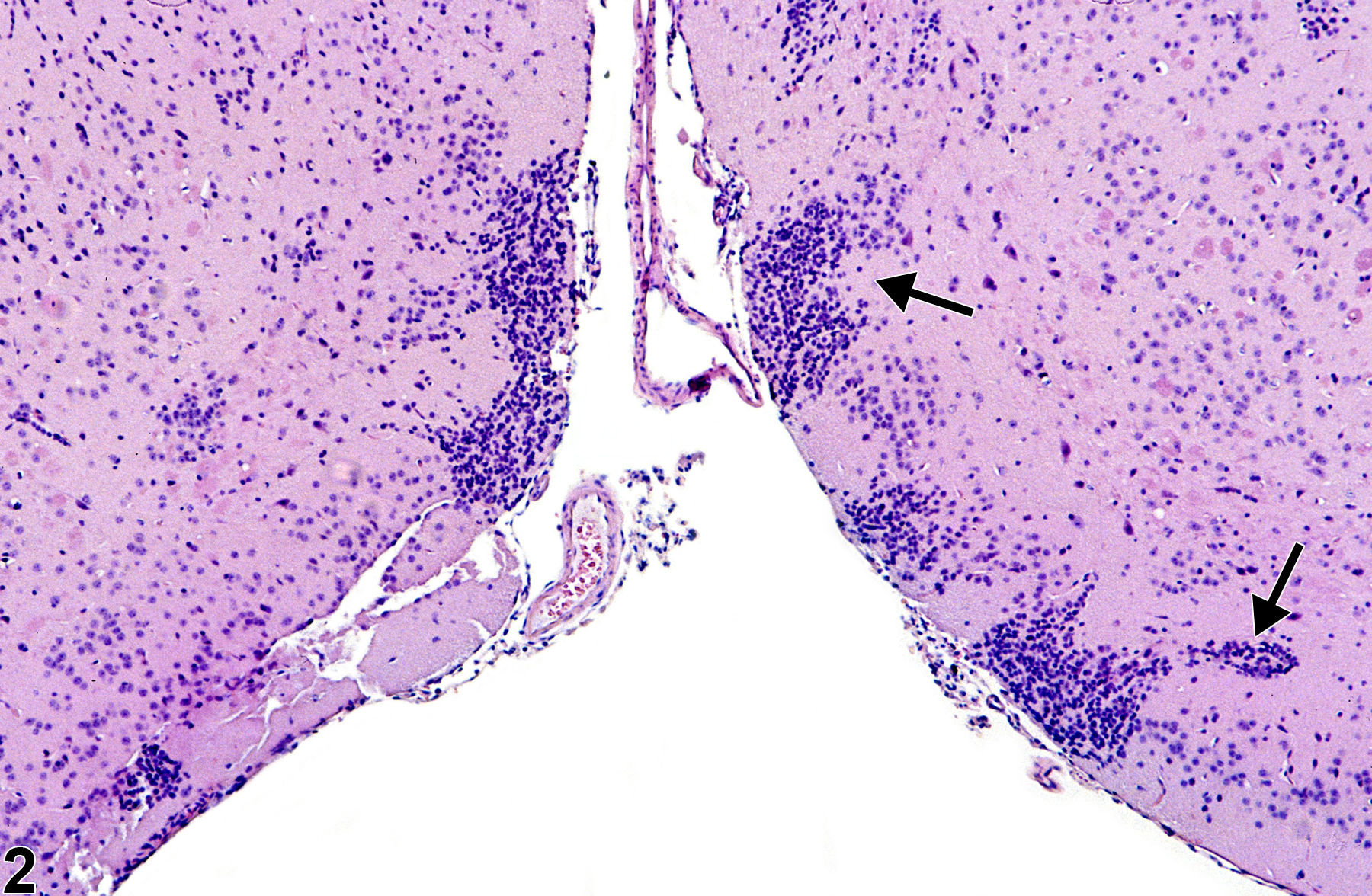
Figure 2. Normal islets of Calleja in a male B6C3F1 mouse from a chronic study. The arrows identify the typical location and morphology of the islets of Calleja.
Figure 2 shows the Islets of Calleja in the frontoparietal cortex. These are a common, normal occurrence of symmetrical clusters of primitive neural cell components in the ventral frontal cortical regions. They are named after C. Calleja, who in 1893 first described these cell aggregations in the olfactory tubercle. They are known to consist of primordial neural cells capable of multiple differentiation pathways to glial and neuronal cells. Many are spongioblasts that may differentiate to either oligodendroglial or astrocytic lineages, and the others are neuroblasts that may differentiate to neurons. In more mature brains, these aggregations are less prominent, having been either utilized in the developing immature brain or effaced by apoptosis that occurs with any unwanted neural cells. Neuroblasts in these locations can be best identified in hematoxylin and eosin sections by a more prominent nucleolus, whereas spongioblasts have a compact nucleus. Increasingly, special immunohistochemical techniques to identify these cells use such antibodies as glial fibrillary acidic protein for astrocytes, Olig-2 for oligodendroglia, and chromogranin, neuron-specific enolase, and so forth, for neurons. Granule neurons in these islets have a high density of D3 (dopamine) receptors and may have less understood neurophysiologic functions. Aggregations of primordial and granule neural cells are frequently misdiagnosed as inflammatory infiltrates. Their location and bilateral symmetry act as helpful indicators of their normal status.
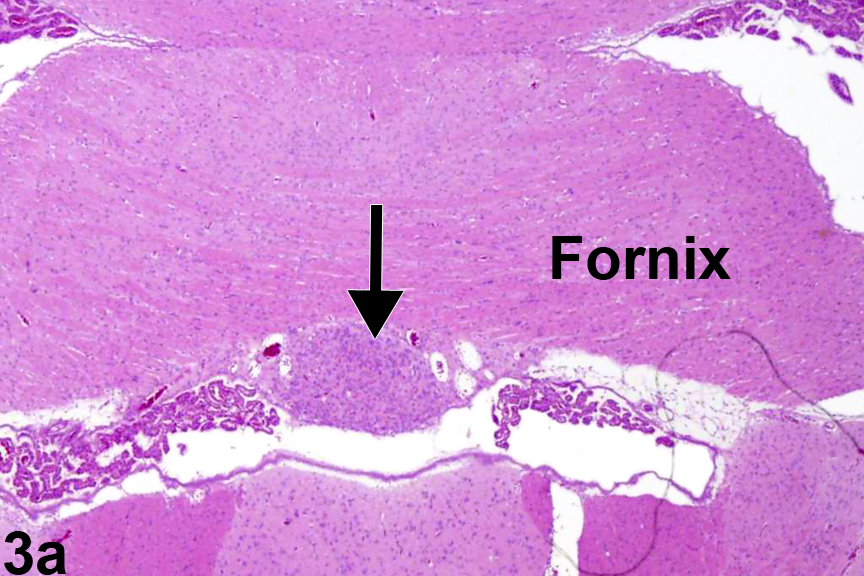
Figure 3a. Normal circumventricular subfornical organ in a male Sprague-Dawley rat at 6 months. The arrow identifies the location of the subfornical organ.
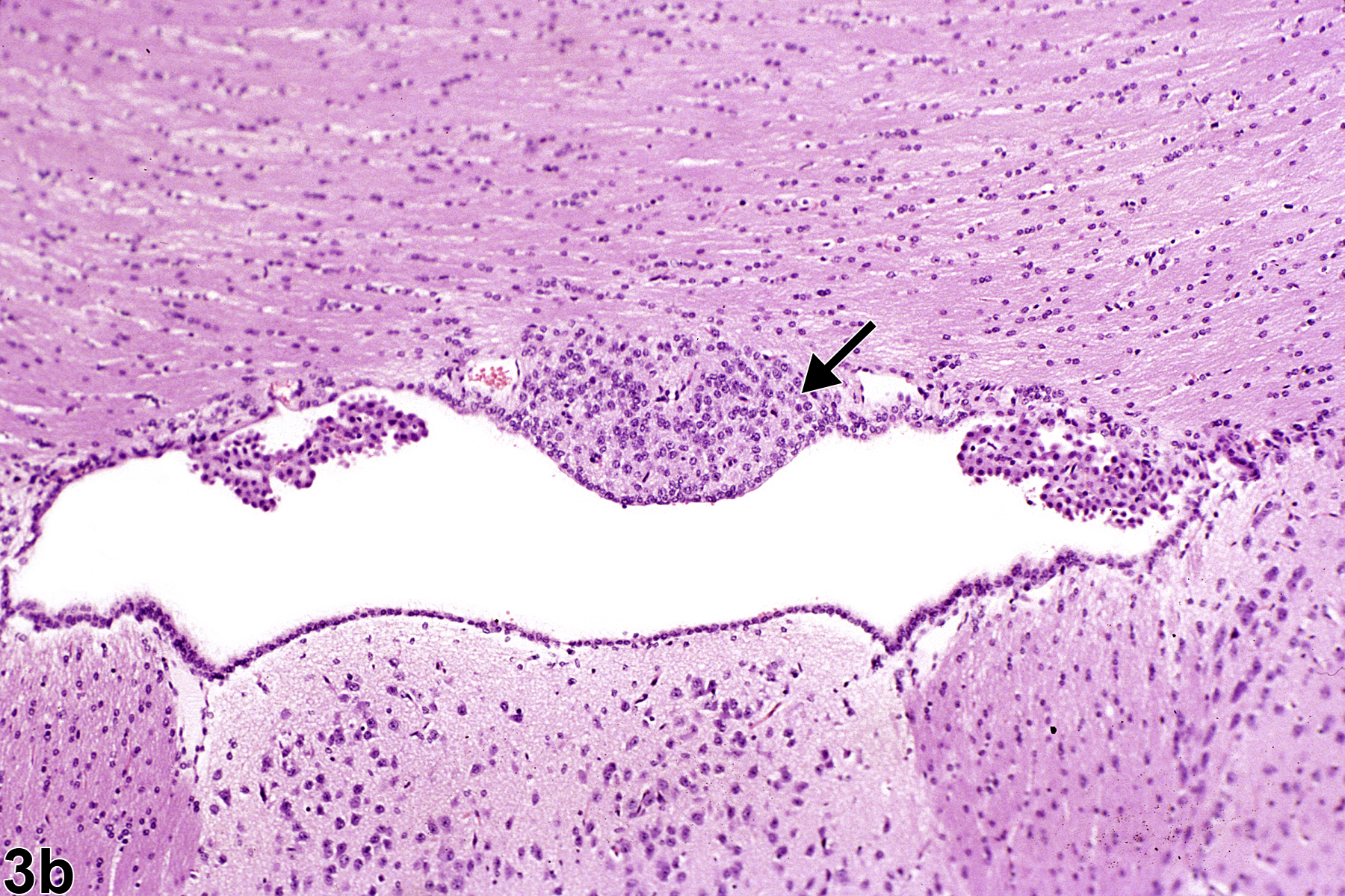
Figure 3b. Normal circumventricular subfornical organ (arrow), at higher magnification, in a male B6C3F1 mouse.
It is important for pathologists to be aware of the various circumventricular organs (CVOs) and their appearance, location, and function. They have in common a lack of the normal blood-brain barrier and may be particularly vulnerable to certain pharmaceutical and toxic compounds that normally cannot penetrate through an effective blood-brain barrier. Adriamycin used as an anticancer agent effectively damages some CVOs. Their vulnerability, because of a lack of blood-brain barrier in these regions, makes their evaluation in toxicity studies particularly important.
CVOs occur at several sites in the brain adjacent to the cerebrospinal fluid pathways (see Table 1). The CVOs are important sites of neural monitoring and system regulation that function by transducing plasma hormonal signals to electrical messages. They are sources of varied neurotransmitters, cytokines, and hormones associated primarily with homeostatic functions involving fluid composition, volume, and osmolality. The cells commonly have a monomorphic appearance within a generally well-vascularized stroma, and some of these structures are commonly misinterpreted as neoplasms, such as neuroblastoma or medulloblastoma. CVOs should not be misdiagnosed as neoplasms.
Images of the subfornical organ and subcommissural organ are included here. Images and description of other CVOs (organum vasculosum of the lamina terminalis, median eminence, pineal, and area postrema) are noted in Garman (2011).
Figure 3a shows the location of the circumventricular subfornical organ, one of seven components of the CVOs. As the name indicates, it is located beneath the fornix at the interventricular foramina (of Monro). The fornix is a densely myelinated tract that connects the hippocampus primarily to the diencephalic structures, thus allowing for neural behavioral influences from the hippocampus to regulate release of hormones from the hypothalamus and pituitary pars distalis. Figure 3b shows, in more detail, the nature of the cells comprising the subfornical organ (arrow) at the juncture of both the lateral and the third ventricle. Notice how it lies immediately below the ependyma, adjacent to the ventricular system, with easy access for dispersion of neurohormones to and from the cerebrospinal fluid. Components of the choroid plexus lie at either side. This unimpressive, monomorphic collection of subfornical organ cells understates their neuronal cell origin and important neurohormonal function(s). The subfornical organ functions in the maintenance of fluid homeostasis by receiving enhanced angiotensin II signaling from plasma for integrating the thirst response to dehydration.
| Circumventricular Organ | Abbreviation | Function | Target | Hormones Involved |
|---|---|---|---|---|
| Subfornical Organ* | SFO | Thirst regulation | Medial preoptic nucleus | Angiotensin II |
| Pineal gland | PI | Circadian rhythms | Subventricular zone | Melatonin |
| Neural lobe, pituitary | NLP | Kidney water transfer; uterine contraction | Kidney tubules; uterine musculature | Antidiuretic hormone; oxytocin |
| Median eminence | ME | Endocrine regulation | Pituitary pars distalis control | Various hormone-releasing factors |
| Organum vasculosum of lamina terminalis* | OVLT | Fluid regulation | Hypothalamic nuclei | |
| Subcommissural organ | SCO | Spinal canal and ventricular development | Spinal cord and ventricles | Spondin |
| Area postrema | AP | Gastric evacuation | Vomition |
*The organum vasculosum of the lamina terminalis and the subfornical organ are interconnected with the medial preoptic nucleus of the hypothalamus, and together they comprise the AV3V region, located anterior and ventral to the third ventricle. This region is important in fluid and electrolyte regulation, controlling thirst, sodium excretion, blood volume regulation, and vasopressin secretion.
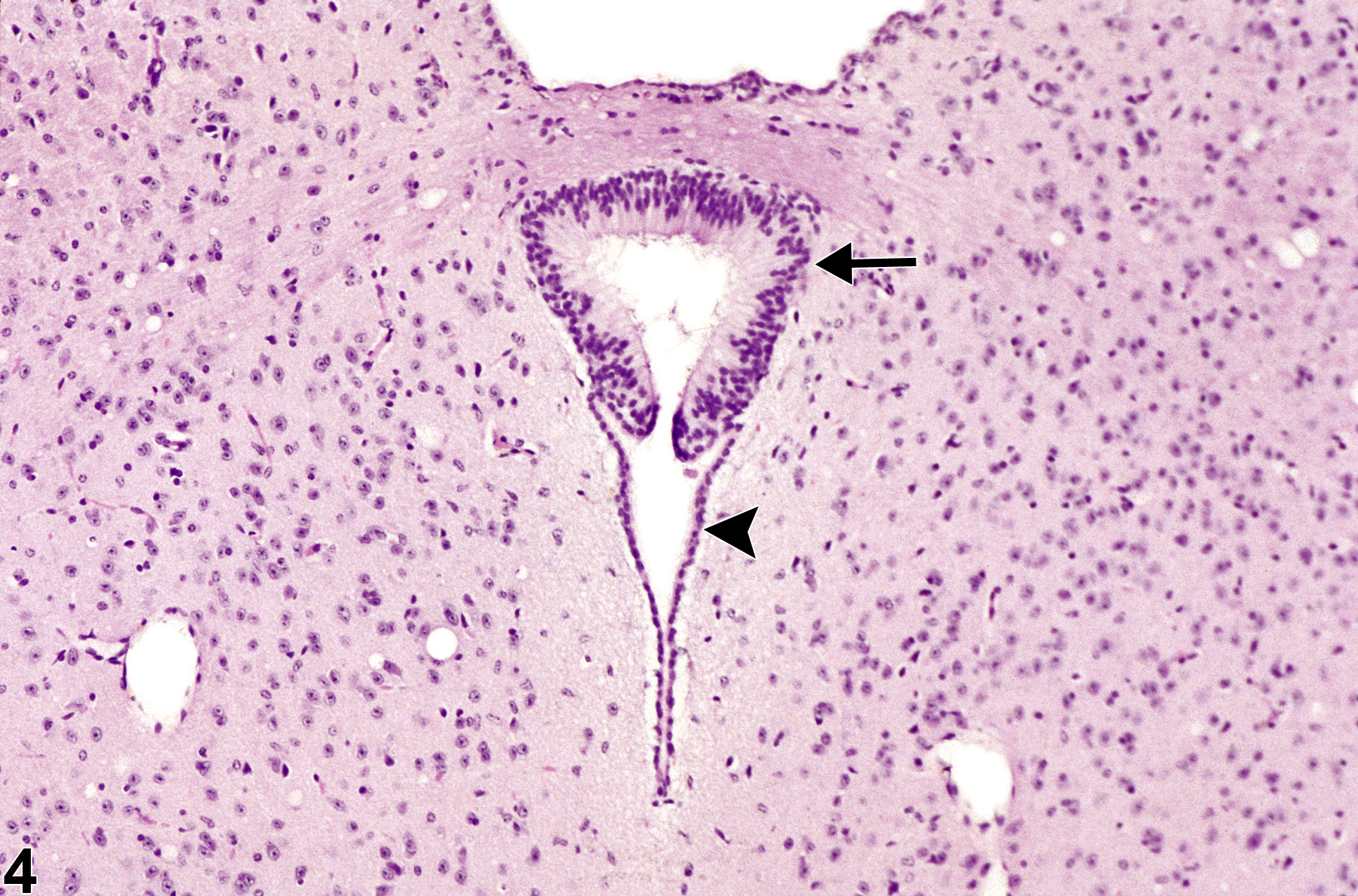
Figure 4. Normal circumventricular subcommissural organ in a female B6C3F1 mouse from a chronic study. Note the prominent columnar appearance of the subcommissural organ cellular components (arrow) compared with the subjacent normal cuboidal ependymal cells of the cerebral aqueduct (arrowhead).
Figure 4 shows the subcommissural organ, another CVO. It is located in the roof of the cerebral aqueduct (aqueduct of Sylvius) that connects the third and fourth ventricles. Note the complexity of the ependymal cells at this site, with cells appearing elongated and pseudostratified (arrow) compared with the subjacent normal cuboidal ependymal cells of the cerebral aqueduct (arrowhead). It is this complex appearance and the focality of the organ that cause some to misdiagnose this structure as a neoplasm, such as an ependymoma. The subcommissural organ is responsible for the formation of Reissner's fiber during spinal cord development and has a role in the neural/adrenal axis and mineralocorticoid regulation. The subcommissural organ should not be misdiagnosed as a neoplasm.
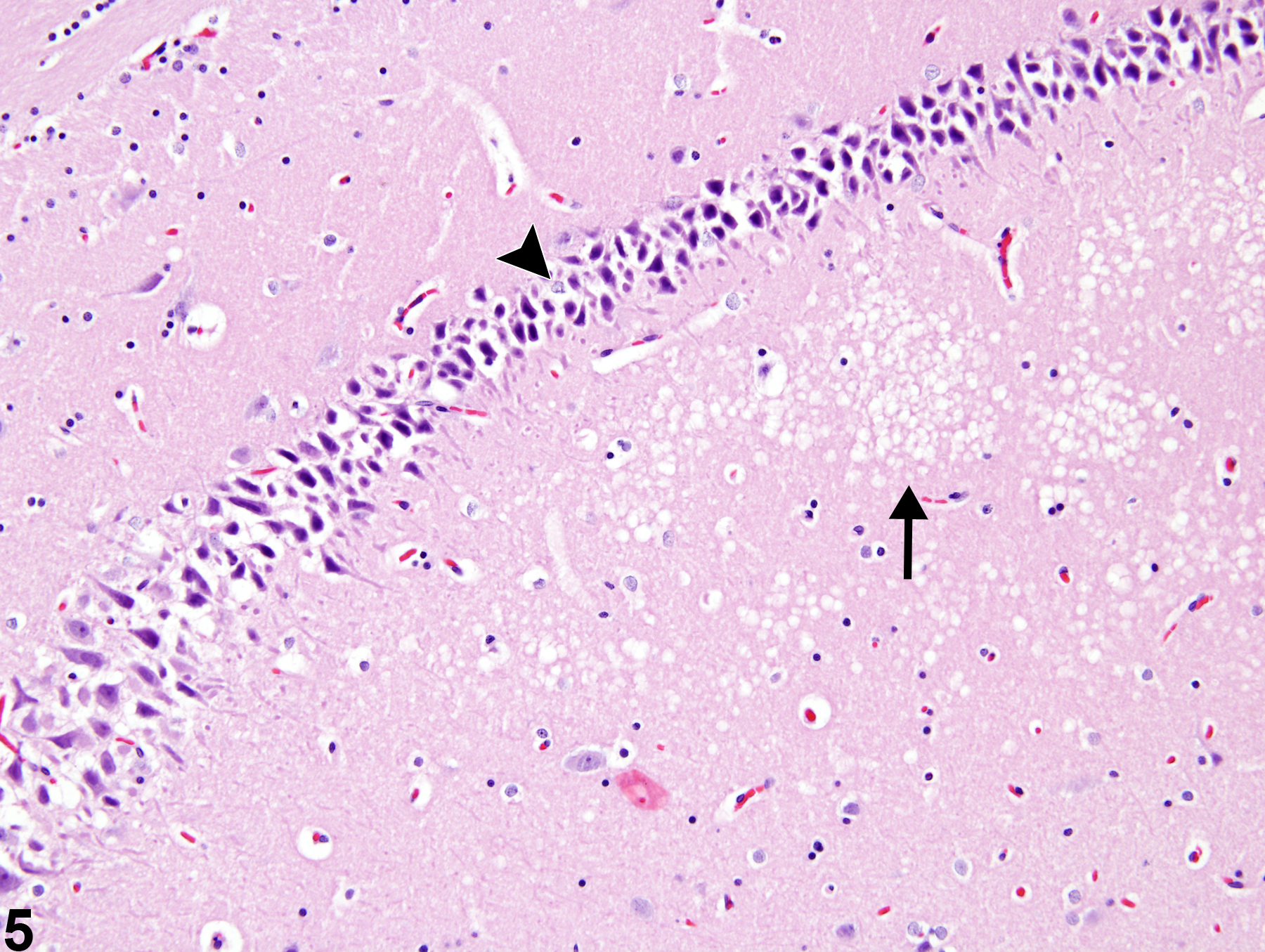
Figure 5. Artifactual basophilic neurons (arrowhead) and artifactual white matter vacuolation (arrow) from a male F344/N rat in a chronic study.
Artifactual findings in the nervous system of rodents are probably the most common and most distracting change that neuropathologists deal with. Figure 5 depicts two common artifacts in the brain: basophilic or dark neurons and white matter vacuolation. These represent basophilic shrinkage and vacuolation of the neuropil, respectively.
Neuronal basophilia and shrinkage are common artifacts in nonperfused brains. They are related to coarse handling and compression of unfixed or poorly fixed brain at the time of dissection from the calvarium and are also the result of inappropriate drying of brain tissue prior to fixation and at the time of fixed brain sectioning. Blocking glutamate receptors in cerebral biopsies has been reported to eliminate the occurrence of dark neuronal artifact. This neuronal change can be found throughout the brain but is most common in cortex, hippocampus, and cerebellar Purkinje cells. Characteristically, there is nuclear and cytoplasmic shrinkage and increased basophilia (Figure 5, arrowhead). Nuclei are shrunken but not karyorrhectic. Genuine antemortem neuronal basophilia, referred to as "chronic nerve cell change," is frequently found adjacent to genuine eosinophilic necrosis of neurons, particularly at the border of ischemic lesions or in lesions induced by other forms of metabolic arrest of neuronal energy metabolism. Numerous toxic compounds have that capability. Authentic basophilic neurons frequently have prominent nuclear basophilic stippling and some evidence of karyorrhexis. The differentiation is often difficult, but in the case of legitimate neuronal basophilia, there are other indicators of neural injury, such as adjacent evidence of eosinophilic neuronal necrosis, capillary endothelial hypertrophy, and the presence of inflammatory infiltrates particularly activated microglial cells (rod cells).
Figure 5 also depicts white matter vacuolation, another important artifact. In this image, artifactual vacuolation of the neuropil (arrow) is subjacent to the basophilic artifact, which makes interpretation of the neuronal basophilic changes even more difficult. White matter vacuolation is a very common postmortem artifact associated with delayed fixation and also improper handling of the tissue during fixation, such as excessive exposure to alcohol in the staining process. Since the nervous system has high lipid content, processing and loss of lipid through solvent extraction for paraffin embedding will normally produce focal to widespread, irregular, fine to large vacuoles predominantly in the white matter (corona radiata of cerebrum, corpus callosum, internal capsule, cerebellar deep white matter), commonly referred to as white matter vacuolation or vacuolar artifact. Artifactual vacuolar change can be induced by holding fixed tissue in 70% alcohol for prolonged periods (48 hours). Such changes often occur in an irregular pattern in densely myelinated structures, whereas genuine vacuolar lesions are more commonly bilaterally symmetrical. In addition, typically artifactual vacuoles have smoothly rounded outlines and are generally empty (i.e., no cellular debris). As a cautionary note, vacuolation is also enhanced in autolyzed tissues and may be accompanied by mild astrocytic swelling and condensation of nuclei accompanying mild neuropil vacuolation in the gray matter. Genuine white matter vacuolation, those lesions due to toxin exposure and alteration of the oligodendroglial axonal myelin wrappings, is commonly bilaterally symmetrical and may have evidence of axonal injury displayed in the affected region as swollen axons or spheroid formation.
Artifactual basophilic neuronal change is significant since its presence may distract the pathologist's attention from the presence of genuine lesions. The pathologist must guard against quickly deciding that all basophilic neuronal change is artifact and recognize that genuine neuronal lesions may accompany this type of artifact. The obvious solution is the use of formalin perfusion fixation followed by 48-72 hours in situ within the calvarium, which is the procedure commonly used in dedicated neurotoxicologic studies. A large number of studies however, for reasons of cost and time expended, still use immersion fixation, and pathologists will therefore continue to face this challenge. Artifactual basophilic neuronal change is not diagnosed in NTP studies.
Artifactual white matter vacuolation is significant since it needs to be differentiated from genuine white matter lesions such as age-associated degenerative changes, intramyelinic edema, axon degeneration, and demyelination. When genuine white matter lesions are present and cannot be differentiated from axon degeneration, demyelination, and intramyelinic edema under light microscopy, the lesion is recorded as "white matter vacuolation" in NTP studies followed by specific evaluation by a neuropathologist.
Balaban CD, Schuerger RJ, Severs WB. 1994. Evidence for a noradrenergic projection to the subcommissural organ. Neurosci Lett 180:209-213.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/7700581Bayer SA. 1987. Neurogenesis in the olfactory tubercle and islands of Calleja in the rat. Int J Dev Neurosci 3(2):135-145.
Abstract: http://www.sciencedirect.com/science/article/pii/0736574885900048Bigotte L, Olsson Y. 1983. Toxic effects of adriamycin on the central nervous system. Ultrastructural changes in some circumventricular organs of the mouse after intravenous administration of the drug. Acta Neuropathol 61:291-299.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/6650142Bolon B, Garman RH, Pardo ID, Jensen K, Sills RC, Roulois A, Radovsky A, Bradley A, Andrews-Jones L, Butt M, Gumprecht L. 2013. STP position paper: Recommended practices for sampling and processing the nervous system (brain, spinal cord, nerve, and eye) during nonclinical general toxicity studies. Toxicol Pathol [Epub ahead of print], doi: 10.1177/0192623312474865.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/23475559Cammermeyer, J. 1961. The importance of avoiding “dark” neurons in experimental neuropathology. Acta Neuropathol 1:245-270.
Abstract: http://link.springer.com/article/10.1007/BF00687191Cammermeyer J. 1978. Is the solitary dark neuron a manifestation of postmortem trauma to the brain inadequately fixed by perfusion? Histochemistry 56:97-115.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/97249Dundore RL, Wurpel JN, Balaban CD, Harrison TS, Keil LC, Seaton JF, Severs WB. 1987. Site-dependent central effects of aldosterone in rats. Brain Res 401:122-131.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/3815089Garman RH. 2011. Histology of the central nervous system. Toxicol Pathol 39:22-35.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/21119051Hooper PT. Finnie JW. 1987. Focal spongy changes in the central nervous system of sheep and cattle. J Comp Pathol 97:433-440.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/3667997Owen GS, Halliwell JV. 2001. Electrophysiological characterization of laminar synaptic inputs to the olfactory tubercle of the rat studied in vitro: Modulation of glutamatergic transmission by cholinergic agents is pathway-specific. Eur J Neurosci 13:1767-1780.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/11359528Johanson C, Stopa E, McMillen P, Roth D, Funk J, Krinke G. 2011. The distributional nexus of choroid plexus to CSF, ependyma and brain: Toxicologic/pathologic phenomena, periventricular destabilization and lesion spread. Toxicol Pathol 39:186-212.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/21189316Jortner BS. 2006. The return of the dark neuron. A histologic artifact complicating contemporary neurotoxicology evaluation. Neurotoxicology 27:628-634.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/16650476Kherani ZS, Auer RN. 2008. Pharmacologic analysis of the mechanism of dark neuron production in cerebral cortex. Acta Neuropathol 116:447-452.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/16650476Morgan DL, Little PB, Herr DW, Moser VC, Collins B, Herbert R, Johnson GA, Maronpot RR, Harry GJ, Sills RC. 2004. Neurotoxicity of carbonyl sulfide in F344 rats following inhalation exposure for up to 12 weeks. Toxicol Appl Pharmacol 200:131-145.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/15476866Radovsky A. Mahler JF. 1999. Nervous system. In: Pathology of the Mouse: Reference and Atlas (Maronpot RR, Boorman GA, Gaul BW, eds). Cache River Press, Vienna, IL, 445-470
Rao DB, Little PB, Malarkey DE, Herbert RA, Sills RC. 2011. Histopathological evaluation of the nervous system in routine National Toxicology Program rodent studies: A modified approach. Toxicol Pathol 39:463-470
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/21430177Rao DB, Little PB, Sills RC. Subsite awareness in neuropathology evaluation in NTP studies: A review of neuroanatomy and functional significance. Toxicol Pathol.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/24135464Summers BA, Cummings JF, de Lahunta A. 1995. Principles of neuropathology. In Veterinary Neuropathology. Mosby, St. Louis, MO, 1-67.
Wells GA, Wells M. 1989. Neuropil vacuolation in brain: A reproducible histological processing artefact. J Comp Pathol 101:355-362.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/2691536