

Nervous System
Brain - Microgliosis
Narrative
The microglia are early-responding cells and are apparent in and around an injured site within 12–24 hours. Their evolution to recognizable macrophages is apparent within 48 hours. As macrophages, they have increased cytoplasm, and products of lipid degradation can be detected with periodic acid Schiff reagent and by autofluorescence at 365 nm wavelength. Such phagocytic cells are referred to as lipid phagocytes or, more commonly, “gitter cells.”
Figure 1 depicts microgliosis adjacent to an inflammatory mononuclear perivascular cuff (arrow). Note the many elongated, irregular nuclei of typical microglial cells. This image depicts the response of brain to injury by the activation of resident microglial cells. These mesodermal cells enter the brain at an early embryological stage of neural development and have macrophage capability. In the inactive state, they are distributed throughout the brain and are difficult to identify because of their nondiscriminate round nuclear character and lack of visible cytoplasm. When they are activated by the presence of brain injury, they become motile and migrate to the site of injury. In the process of migration, nuclei become elongated and irregular in shape and are referred to as “rod cells.” By this nuclear morphology, they are easily detected (as shown in this image), distributed distant from the perivascular monocytic infiltrate. As well as resident microglia, mononuclear cells of blood and bone marrow origin cross the endothelium and migrate to the injured site, where they also take up phagocytic functions like those of resident microglia. This accounts for the presence of some degree of perivascular cuffing by monocytes adjacent to injured brain tissue, as shown in Figure 1 (arrow). The appearance and function of these blood borne monocytes are not known to be different from those of resident microglia. A particularly useful immunohistochemical stain to detect these cells in brain is the Iba-1 (ionized calcium binding adaptor molecule 1) antibody.
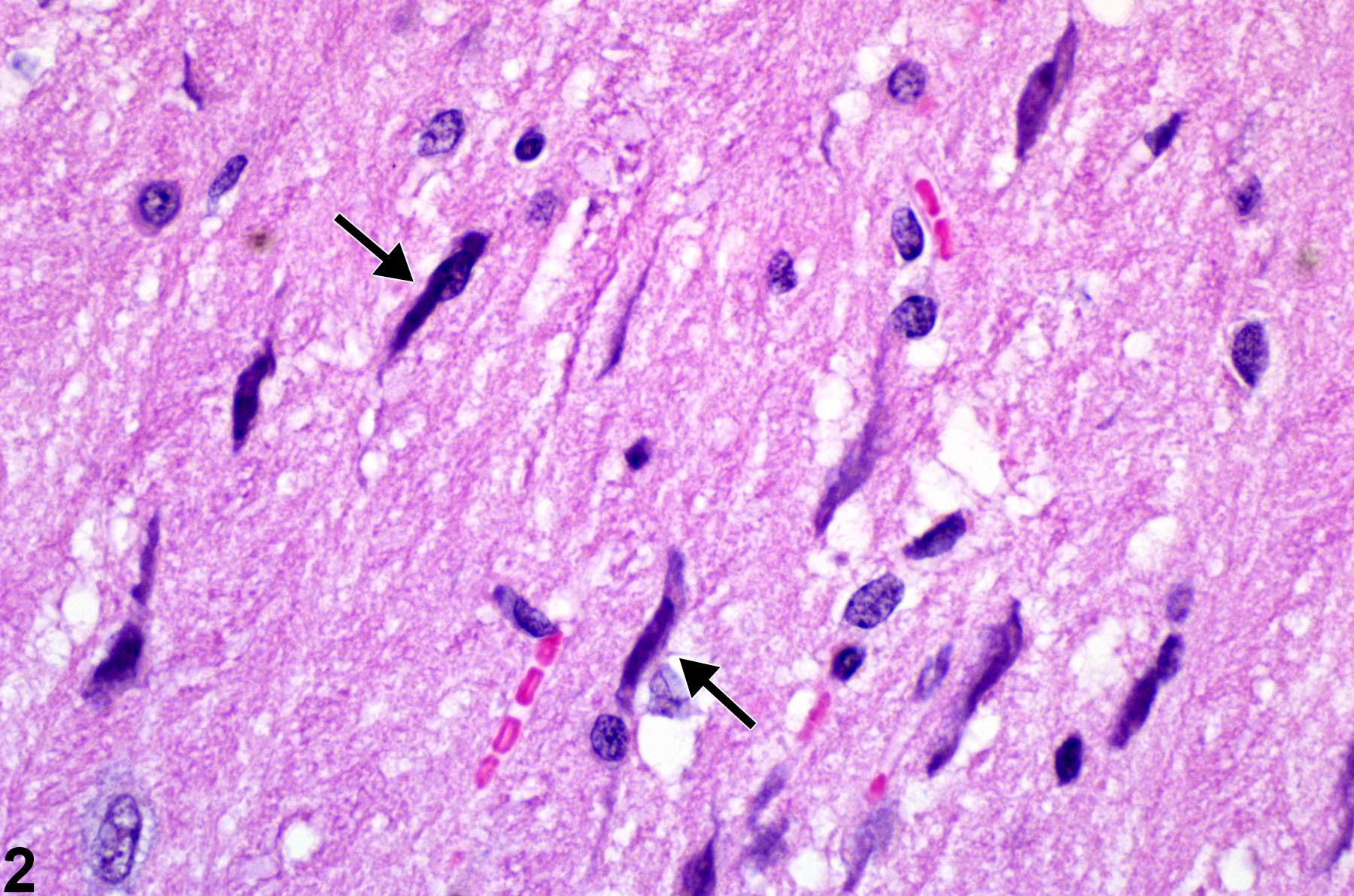
Figure 2 shows activated elongated irregular nuclei (arrows) of microglial cells (rod cells). Shown at high magnification, these activated microglial cells are migrating under chemotactic stimuli to a region of neural injury. It is not possible to say whether these cells are resident microglia or those derived from infiltrating monocytic cells carried to the nervous tissue by the blood stream.
{kind=link}
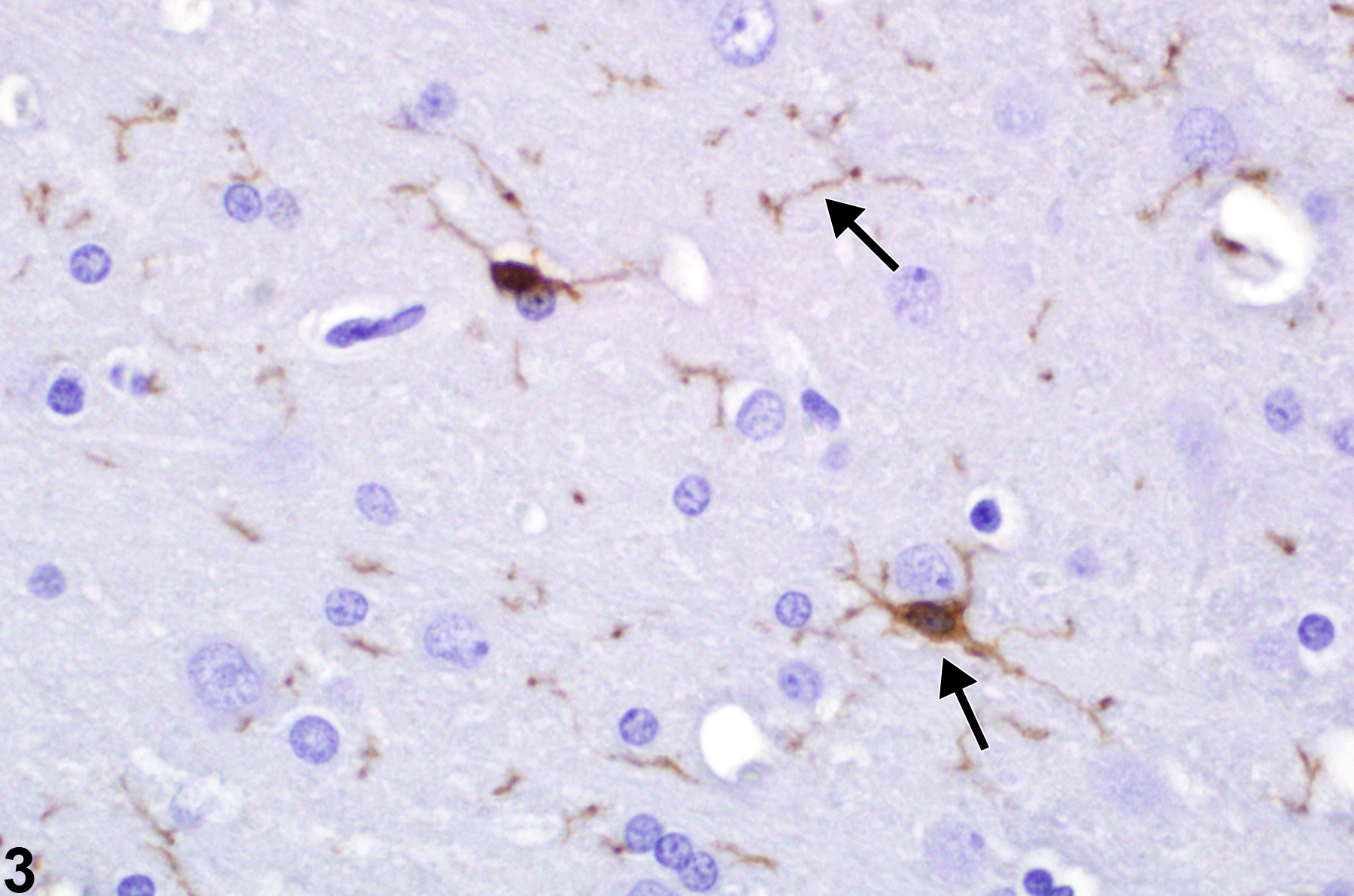
Depicted in Figure 3 is the Iba-1 immunohistochemical detection of microglia in cerebral cortex. Note the brown-staining cells and extensive radiating, cross, and longitudinal sections of their processes (arrows). Iba-1 is specifically expressed in macrophages and microglia and is upregulated during cellular activation.
{kind=link}
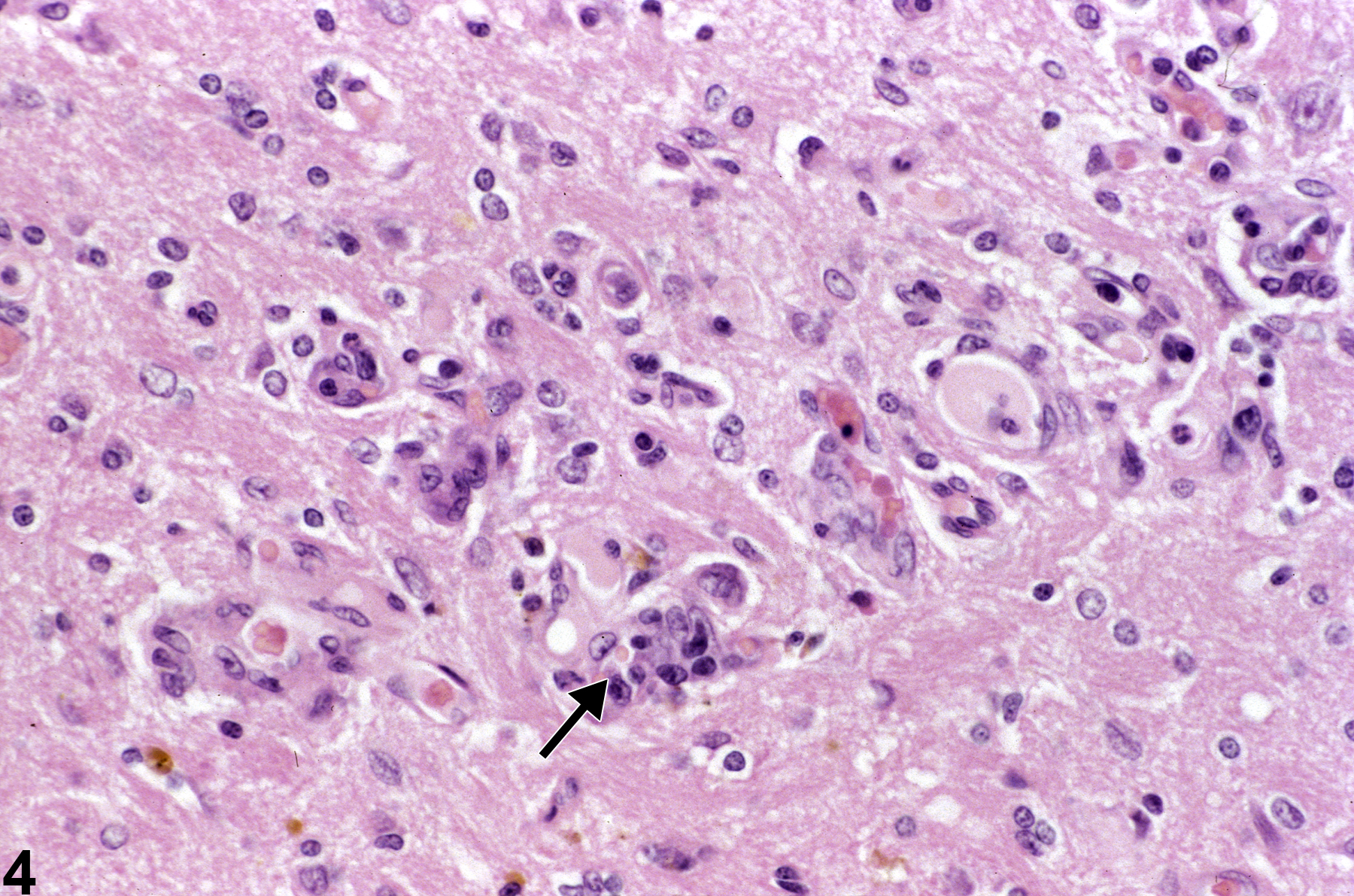
Figure 4 shows reactive gliosis and capillary hyperplasia (arrow) in a site of former injury. Capillaries are quickly responsive to injury of adjacent tissue, and endothelial nuclear hypertrophy is evident within 24 hours. To the pathologist these capillary changes are often, upon low-magnification scanning of brain tissue, the first indication of damage in a region. The reactive gliosis seen in this image is likely a mixture of infiltrating blood monocytes and resident microglial cells. None have yet differentiated to recognizable macrophages. The lesion is therefore probably less than 48 hours old. Selective study of endothelial responses may be enhanced by the use of immunohistochemistry using laminin and von Willebrand factor.
{kind=link}
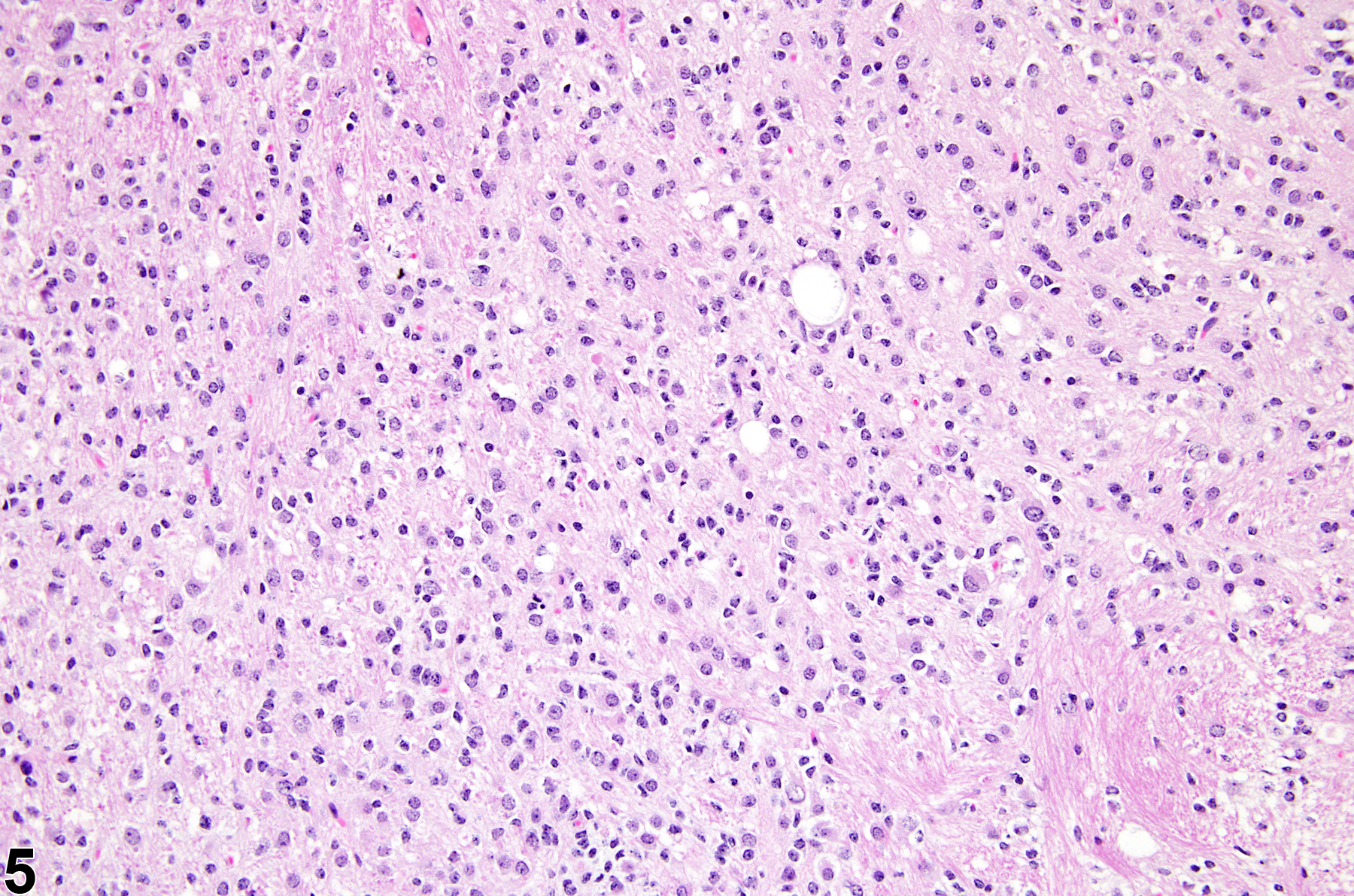
Figure 5 displays a mixed gliotic response to injury of the putamen, consisting of reactive astrocytes, macrophages, microglia, and mononuclear cells. It is apparent that there is a significant increase in nucleated cells in the region. At this magnification, and with hematoxylin and eosin (H&E) staining alone, it is not possible to identify the specific cell types that make up the population. In many studies, it is only relevant to identify the response as being of mixed glial type, which would include astrocytes, microglial cells, and monocytic cells of blood origin and/or macrophages if they have already differentiated to phagocytic cells. In those cases where specific identification of the types of infiltrating cells is necessary, special stains and immunohistochemistry would be required.
{kind=link}
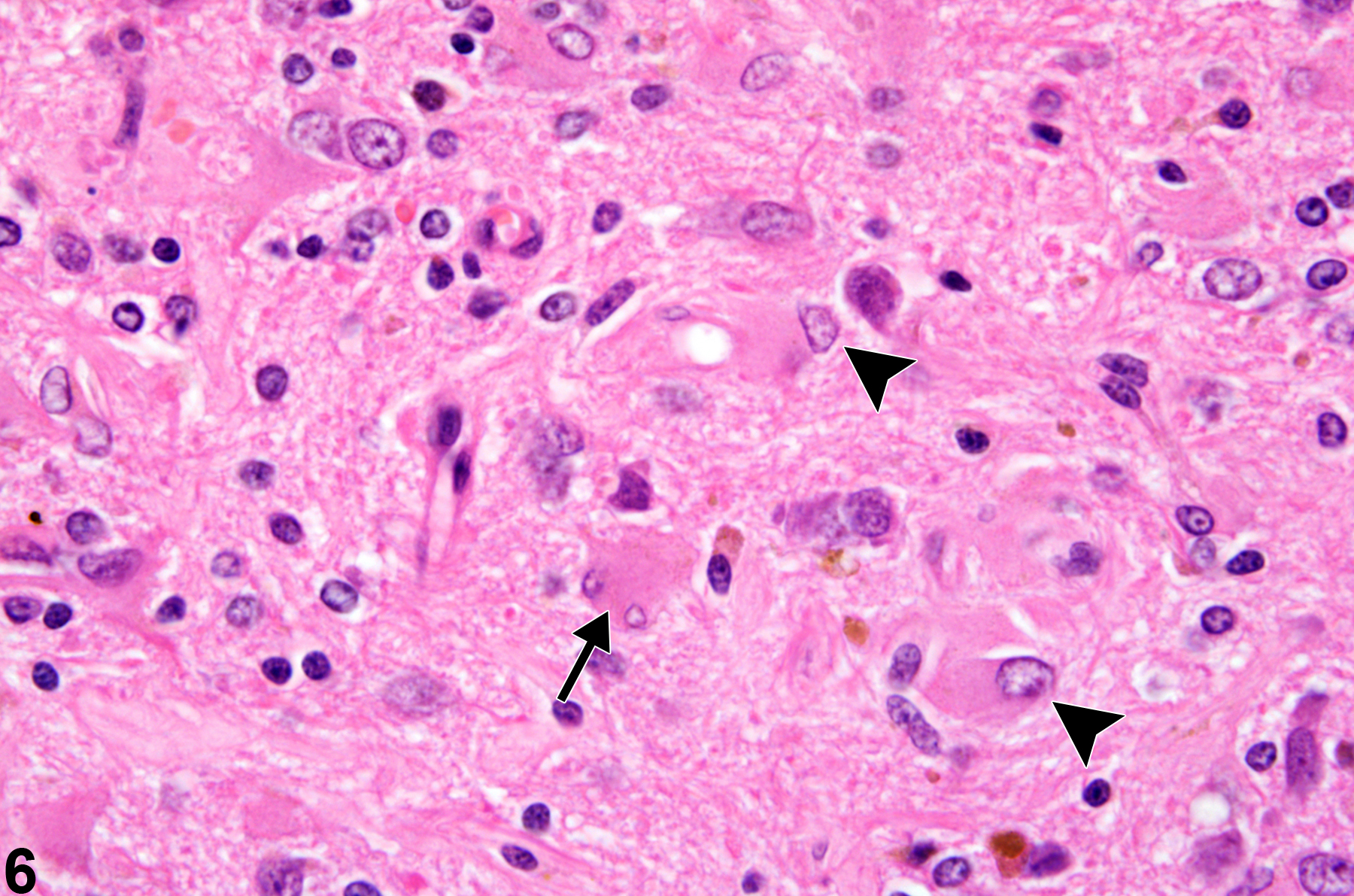
Normal astrocytes, protoplasmic and fibrillary, are diffusely distributed in the nervous system, where they serve, among other functions, to stabilize structure and maintain the neural milieu, neuronal energy processes, and the blood brain barrier. The density of the normal astrocyte population in the brain is increased in the glia limitans just beneath the pia mater, and also around blood vessels.Gemistocytic astrocytosis is an integral part of the progressive response to many forms of neural injury and is helpful in uncovering subtle brain injury, as well as giving an approximation of the chronology of the lesions present. Figure 6 shows at high magnification reactive gemistocytic astrocytosis in a site of healing neural injury stained with H&E. Note the typical eccentric nucleus and prominent eosinophilic cytoplasm of these reactive astrocytes (arrowheads) and the less frequent micro-binucleate form (arrow). In addition, the normal radiating processes become enlarged and extensive. In this example, there is a rare form of binucleation of one of the reactive astrocytes, signaling the presence not only of hypertrophy but also of hyperplasia in the response. The appearance of gemistocytic astrocytes is a useful indicator of the age of a brain lesion since it is seldom well developed on H&E sections until 12–14 days after injury. The use of the immunohistochemical technique for glial fibrillary acidic protein (GFAP), which constitutes the astrocytic intermediate filaments, will detect the earliest stages of the response. This commences at approximately 7 days postinjury and makes any site of injury more recognizable even if there is little to see with H&E sections. In this way, GFAP is a very useful, sensitive indicator of subtle brain injury and the associated astrocytic responses. While GFAP immunoreactivity is used extensively in neuropathology to identify and quantify astrocytes, some other cell types such as ependymal cells may also express a low level of this intermediate filament.
{kind=link}
In the presence of concurrent lesions, lesions with the most severity are typically diagnosed. Other concurrent lesions may be diagnosed separately, if warranted by the severity.
Fix AS, Ross JF, Stitzel SR, Switzer RC. 1996. Integrated evaluation of central nervous system lesions: Stains for neurons, astrocytes, and microglia reveal the spatial and temporal features of MK-801-induced neuronal necrosis in the rat cerebral cortex. Toxicol Pathol 24:291-304.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/8736385Kimelberg HK. 2010. Functions of mature mammalian astrocytes: A current view. Neuroscientist 16:79-106.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/20236950Sidoryk-Wegrzynowicz M, Wegrzynowicz M, Lee E, Bowman AB, Aschner M. 2011. Role of astrocytes in brain function and disease. Toxicol Pathol 39:115-123.
Abstract: http://www.ncbi.nlm.nih.gov/pubmed/21075920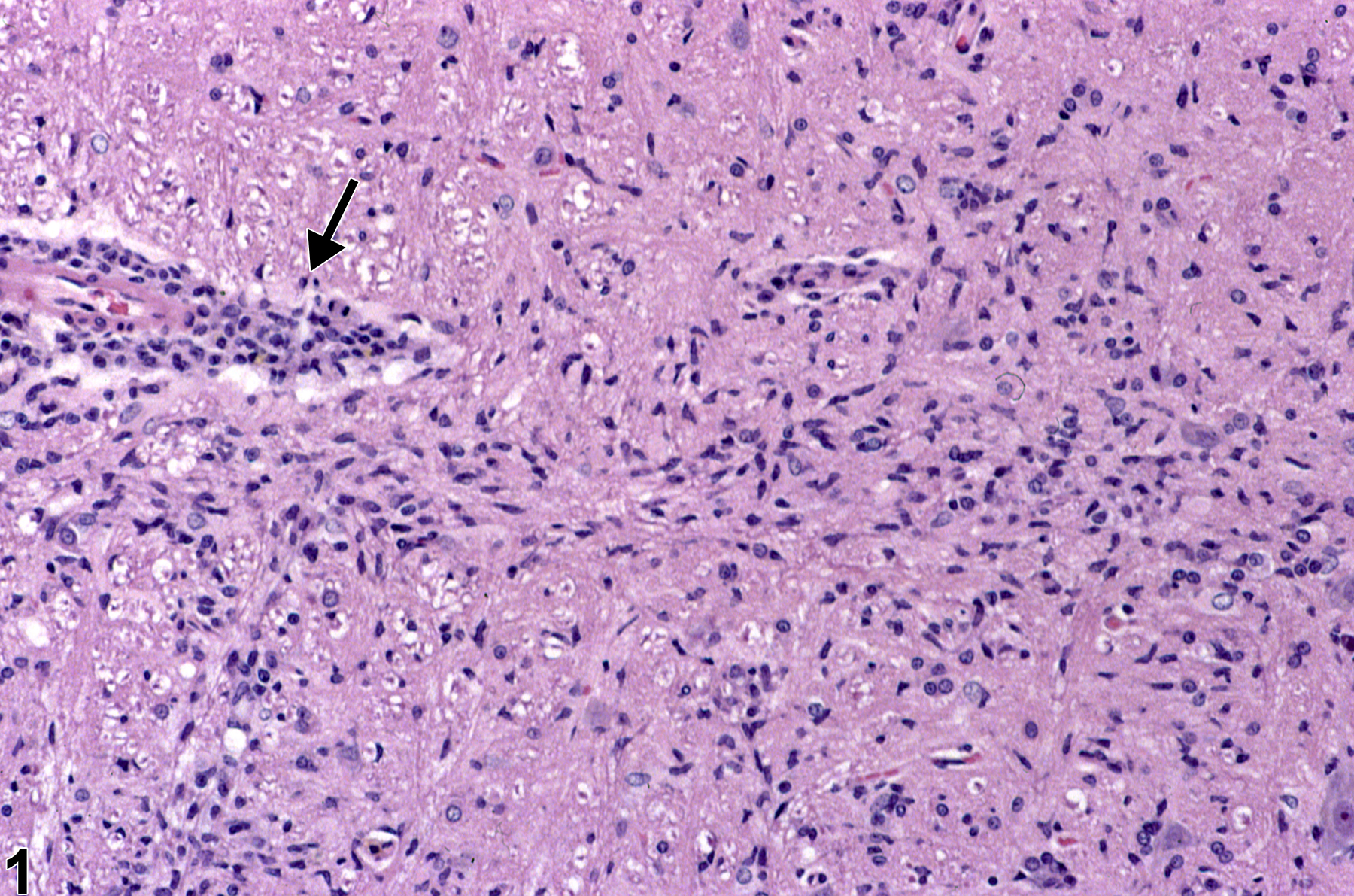
Microgliosis adjacent to an inflammatory mononuclear perivascular cuff (arrow) in a female F344/N rat from a chronic study. Note the many elongated, irregular nuclei of typical microglial cells.
All Images

Microgliosis adjacent to an inflammatory mononuclear perivascular cuff (arrow) in a female F344/N rat from a chronic study. Note the many elongated, irregular nuclei of typical microglial cells.

Nuclei of microglia at a higher magnification in a female F344/N rat from a chronic study. Their elongated irregular nuclear morphology (arrows) is apparent.

A particularly useful immunohistochemical stain using the Iba-1 antibody to detect these cells (arrows) in the brain in an F344/N rat. Other immunohistochemical methods used as microglial markers include OX-42 and CD68.

Reactive gliosis and capillary hyperplasia (arrow) in a site of former injury in a female F344/N rat from a chronic study. Capillaries are quickly responsive to injury of adjacent tissue, and endothelial nuclear hypertrophy is evident within 24 hours.

A mixed gliotic response to injury of the putamen, consisting of reactive astrocytes, macrophages, microglia, and mononuclear cells, in a male B6C3F1 mouse from a chronic study. It is apparent that there is a significant increase in nucleated cells in the region.

Reactive gemistocytic astrocytosis in a site of healing neural injury stained with hematoxylin and eosin, in a treated female F344/N rat from a chronic study. Note the typical eccentric nucleus and prominent eosinophilic cytoplasm of these reactive astrocytes (arrowheads) and the less frequent micro-binucleate form (arrow).