

Endocrine System
Adrenal Gland, Cortex, X-Zone - Atrophy
Narrative
{kind=link}
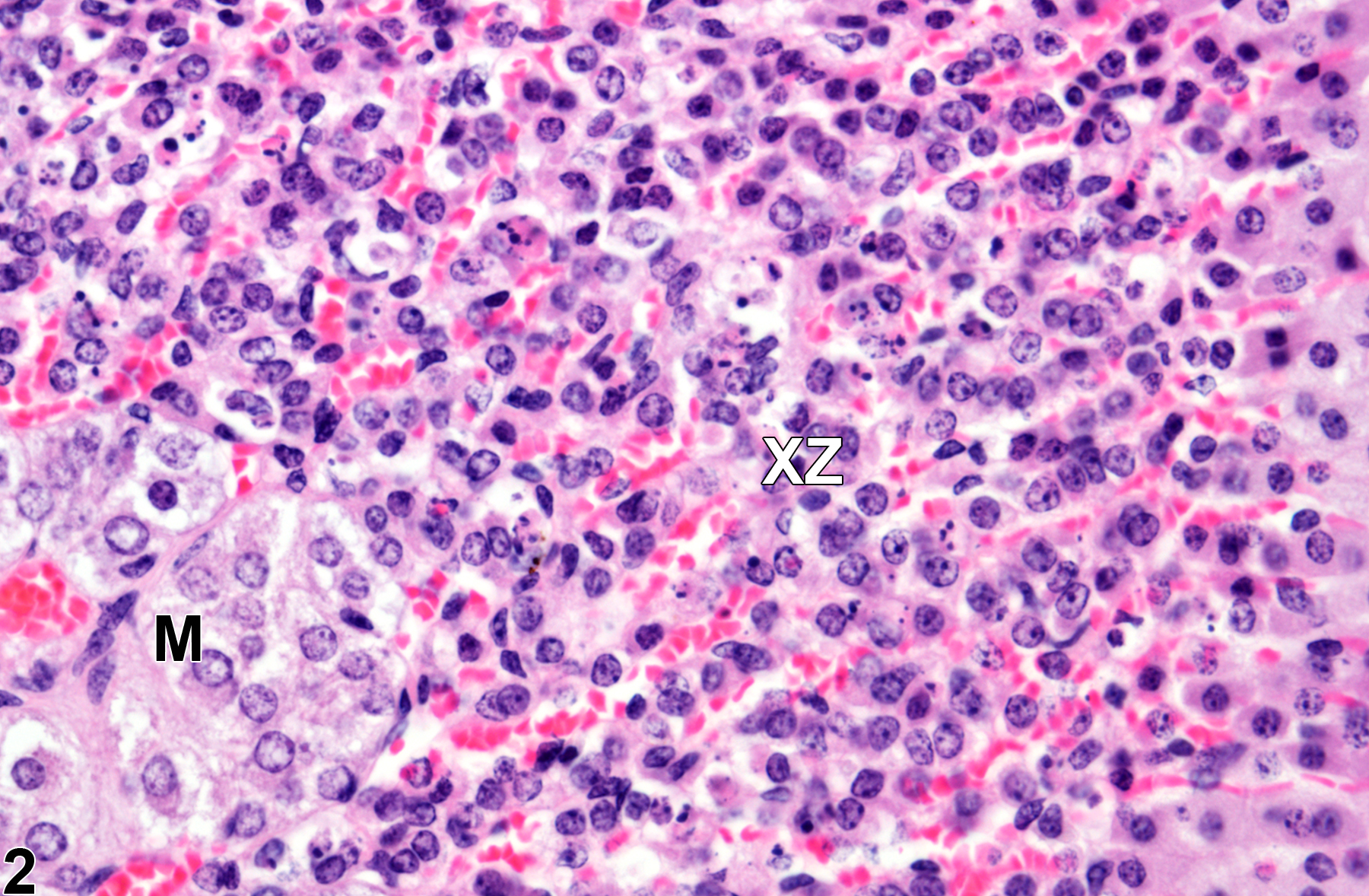
Normal regression (involution) of the X-zone in females of many mouse strains, including the B6C3F1 strain, progresses in morphologically distinct stages. The onset of regression begins with vacuolization of scattered constituent cells (Figure 1 and Figure 2). As regression continues, the number of vacuolated cells progressively increases until virtually all X-zone cells are affected. In later stages, the vacuolated X-zone cells undergo degeneration and necrosis, with subsequent overall X-zone architectural collapse, condensation, and eventual disappearance. A common end-stage sequela is the residual accumulation of pigment-laden cells in the perimedullary area formerly occupied by the X-zone. In males, X-zone regression is similar except that it usually occurs without vacuolization.
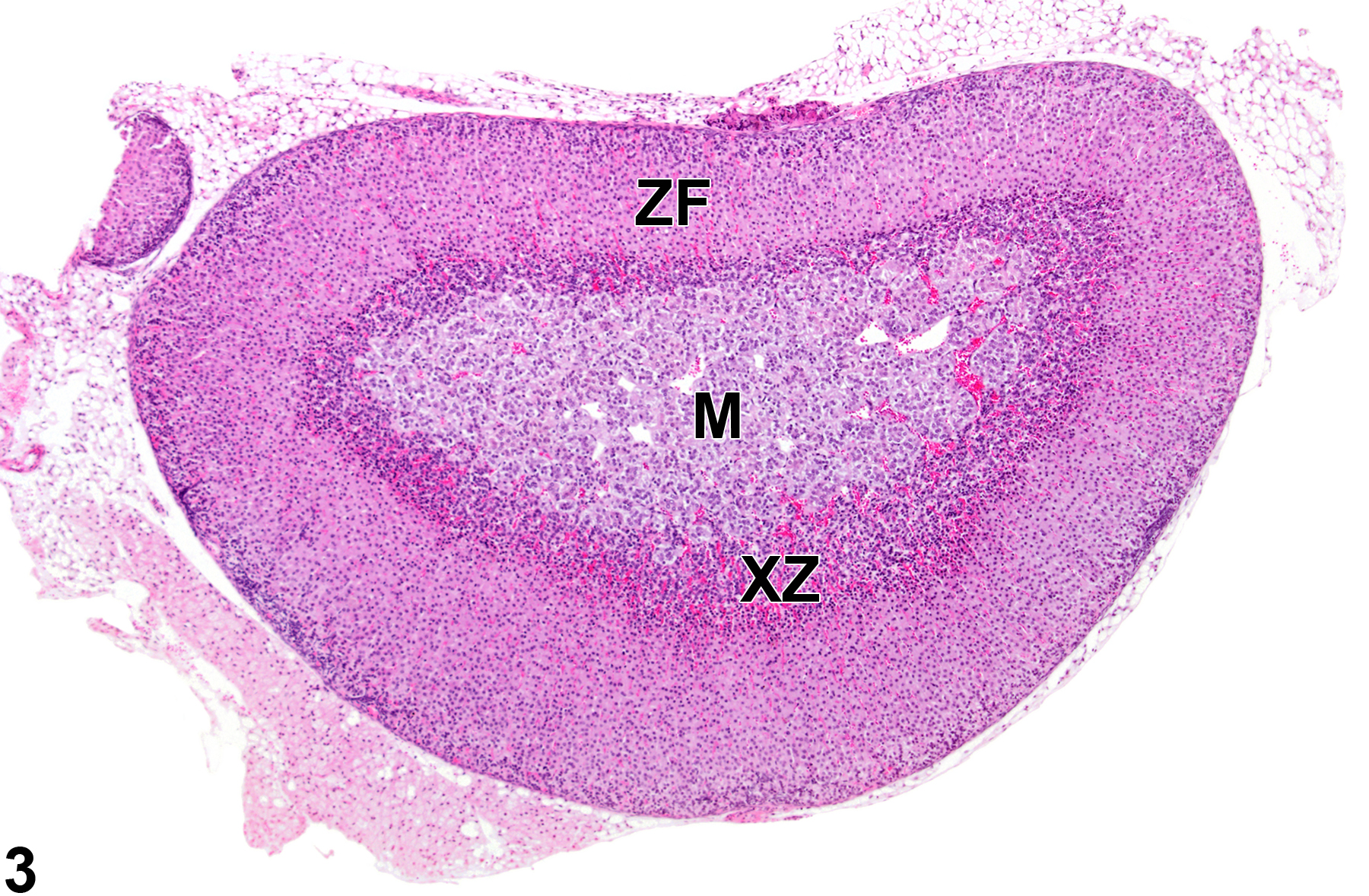
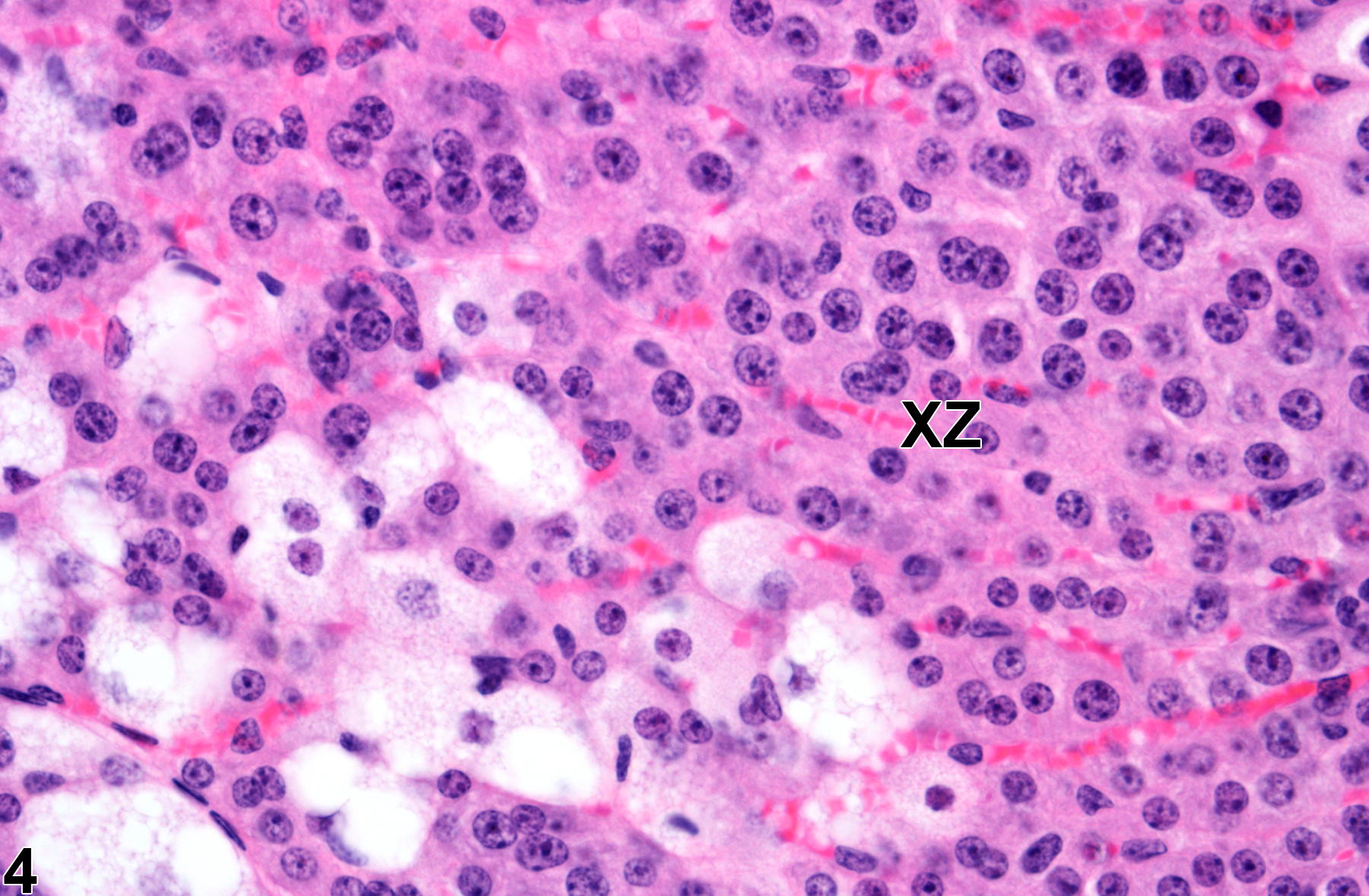
The function of the X-zone is unknown. Its normal development and regression are mediated by gonadal and thyroid hormones, so factors that alter levels of these hormones can affect the X-zone. For example, gonadectomy prolongs the persistence of the X-zone in female mice and prepubertal male mice and can cause the reappearance of the X-zone in postpubertal males. Administration of androgens like testosterone is followed by rapid disappearance of the X-zone in female mice. Administration of certain other chemicals can also affect the X-zone, resulting in asynchronous deviations, such as accelerated regression (Figure 3 and Figure 4) in treated groups compared with age-matched concurrent controls (Figure 1 and Figure 2).
{kind=link}
{kind=link}
Blystone CR, Elmore SA, Will KL, Malarkey DE, Foster PMD. 2011. Toxicity and carcinogenicity of androstenedione in F344/N rats and B6C3F1 mice. Food Chem Toxicol 49:2116-2124.
Abstract: https://www.ncbi.nlm.nih.gov/pubmed/21651954Chhabra RS, Elwell MR, Chou B, Miller RA, Renne RA. 1990. Subchronic toxicity of tetrahydrofuran vapors in rats and mice. Fund Appl Toxicol 14:338-345.
Abstract: https://www.ncbi.nlm.nih.gov/pubmed/2318358Dunn TB. 1970. Normal and pathologic anatomy of the adrenal gland of the mouse, including neoplasms. J Natl Cancer Inst 44:1323-1389.
Abstract: http://jnci.oxfordjournals.org/content/44/6/1323.abstractHershkovitz L, Beuschlein F, Klammer S, Krup M, Weinstein Y. 2007. Adrenal 20α-hydroxysteroid dehydrogenase in the mouse catabolizes progesterone and 11-deoxycorticosterone and is restricted to the X-zone. Endocrinology 148:967-988.
Full Text: https://doi.org/10.1210/en.2006-1100Matsuura S, Suzuki K. 1986. Morphological changes in the submandibular glands and in the X zone of the adrenal gland following ovariectomy in mice. Cell Tissue Res 246:549-556.
Abstract: https://www.ncbi.nlm.nih.gov/pubmed/3791382National Toxicology Program. 1993. NTP TR-443 Toxicology and Carcinogenesis Studies of Oxazepam (CAS No. 604-75-1) in Swiss-Webster and B6C3F1 Mice (Feed Studies). NTP, Research Triangle Park, NC.
Abstract: https://ntp.niehs.nih.gov/go/6030National Toxicology Program. 1996. NTP TR-447. Toxicology and Carcinogenesis Studies of Acetonitrile (CAS No. 79-05-8) in F344/N Rats and B6C3F1 Mice (Inhalation Studies). NTP, Research Triangle Park, NC.
Abstract: https://ntp.niehs.nih.gov/go/6038National Toxicology Program. 2010. NTP TR-560. Toxicology and Carcinogenesis Studies of Androstenedione (CAS No. 63-05-8) in F344/N Rats and B6C3F1 Mice (Gavage Studies). NTP, Research Triangle Park, NC.
Abstract: https://ntp.niehs.nih.gov/go/33555Nyska A, Maronpot RR. 1990. Adrenal gland. In: Pathology of the Mouse: Reference and Atlas (Maronpot RR, Boorman GA, Gaul BW, eds). Cache River Press, Vienna, IL, 509-536.
Rosol TJ, Yarrington JT, Latendresse J, Capen CC. 2001. Adrenal gland: Structure, function, and mechanisms of toxicity. Toxicol Pathol 29:41-48.
Abstract: https://www.ncbi.nlm.nih.gov/pubmed/11215683Shire JGM, Beamer WG. 1984. Adrenal changes in genetically hypothyroid mice. J Endocrinol 102:277-280.
Abstract: https://www.ncbi.nlm.nih.gov/pubmed/6481284Spencer PJ, Crissman JW, Stoot WT, Corley RA, Cieszlak FS, Schumann AM, Hardisty JF. 2002. Propylene glycol monomethyl ether (PGME): Inhalation toxicity and carcinogenicity in Fischer 344 rats and B6C3F1 mice. Toxicol Pathol 30:570-579.
Full Text: http://tpx.sagepub.com/content/30/5/570.full.pdfTanaka S, Matsuzawa A. 1995. Comparison of adrenocortical zonation in C57BL/6J and DDD mice. Exp Anim 44:285-291.
Abstract: https://www.ncbi.nlm.nih.gov/pubmed/8575542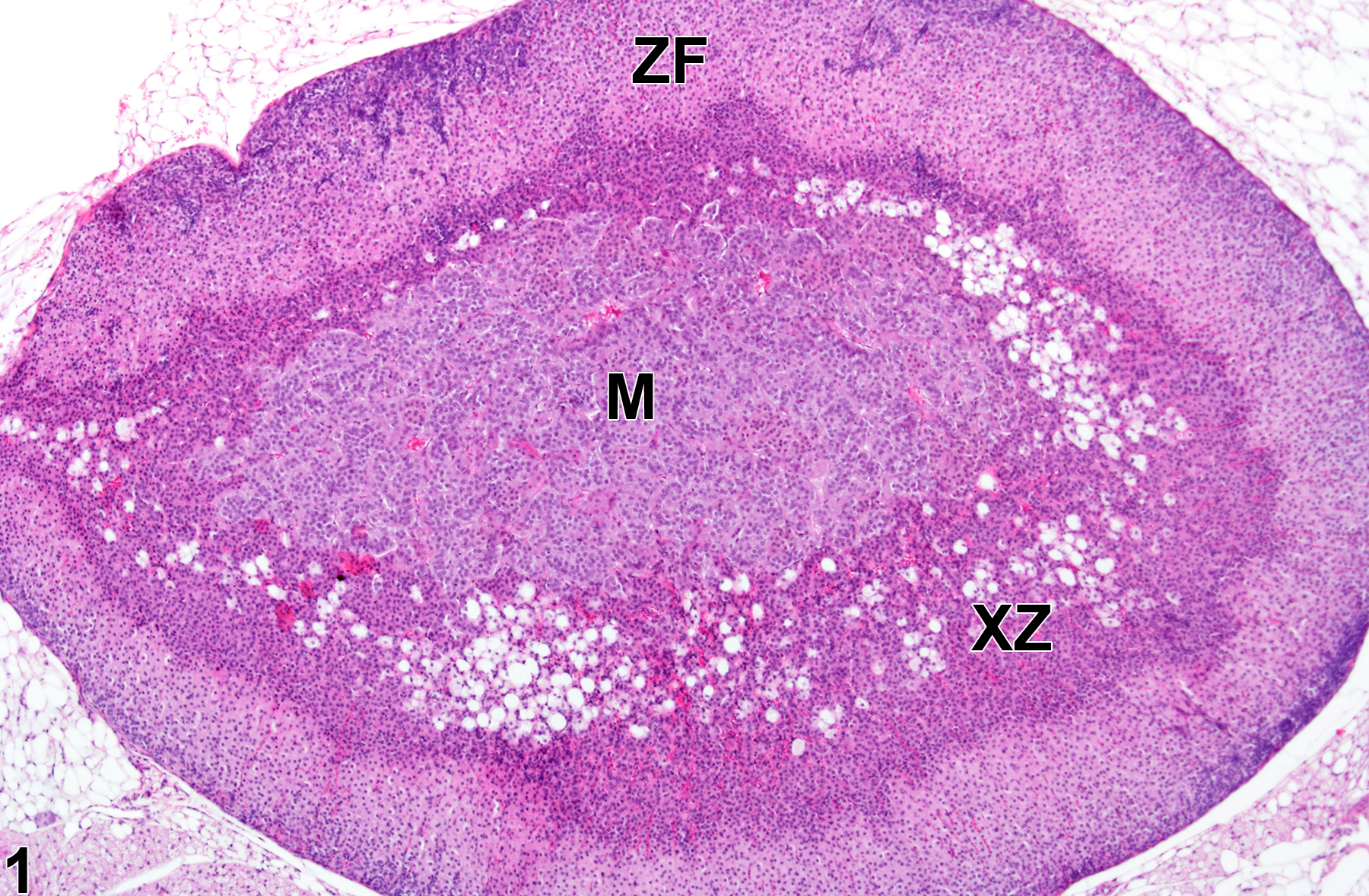
Adrenal gland, Cortex, X-zone - Normal in a female B6C3F1/N mouse from a subchronic study. Adrenal gland with moderate numbers of vacuolated cells in an age-matched control virgin female is shown for comparison with Figure 3. M = medulla, XZ = X-zone, ZF = zona fasciculata.
All Images

Adrenal gland, Cortex, X-zone - Normal in a female B6C3F1/N mouse from a subchronic study. Adrenal gland with moderate numbers of vacuolated cells in an age-matched control virgin female is shown for comparison with Figure 3. M = medulla, XZ = X-zone, ZF = zona fasciculata.

Adrenal gland, Cortex, X-zone - Normal in a female B6C3F1/N mouse from a subchronic study (higher magnification of Figure 1). Adrenal gland in an age-matched control virgin female is shown for comparison with Figure 4. M = medulla; XZ = X-zone.

Adrenal gland, Cortex, X-zone - Atrophy in a virgin female B6C3F1/N mouse from a subchronic study. There is acceleration of X-zone (XZ) regression characterized by narrowing of the X-zone compared with Figure 1. M = medulla, ZF = zona fasciculata.

Adrenal gland, Cortex, X-zone - Atrophy in a virgin female B6C3F1/N mouse from a subchronic study (higher magnification of Figure 3). There is increased degeneration and necrosis of the X-zone (XZ) cells compared with Figure 2.